Received: Mon 26, Jun 2023
Accepted: Wed 19, Jul 2023
Abstract
A-kinase anchoring protein 79/150 (AKAP79/150) is an important scaffolding protein, which anchors various important proteins to locate in appropriate synaptic domains to regulate excitatory synaptic intensity. With sweeping advances in the biology of AKAP79/150 and its critical role in the pathophysiology of various human disorders, more and more evidence is breeding new opportunities for potential therapeutic intervention in an attempt to improve the clinical outcomes of those patients. Herein, we review the basic structure and main functions of AKAP79/150, focusing on the pathophysiological mechanisms of AKAP79/150 in different human disorders, with a particular emphasis on the inflammatory pain, epilepsy, and depression, to discuss their potential therapeutic intervention value in patients with those diseases.
Keywords
AKAP79/150, structure and biological functions, inflammatory pain, epilepsy, depression, potential therapeutics
1. Introduction
AKAP79 is a prototypic A-kinase-anchoring protein (AKAP), with three orthologs (comprising human AKAP79, rodent AKAP150, and bovine AKAP75, gene name AKAP5), which fulfills key physiological roles. As signal-organizing molecules, AKAPs tether those noncatalytic regulatory proteins, which profoundly influence the action of protein kinases and phosphatases, in subcellular environments to control the phosphorylation state of neighboring substrates [1].
AKAP79/150 directs its cohort of anchored enzymes toward selected transmembrane proteins to facilitate their efficient regulation [1]. For example, AKAP79/150 connects with N-methyl-D-aspartate receptors (NMDARs) and α-Amino-3-hydroxy-5-methyl-4-isoxazolepropionic acid receptors (AMPARs) through two membrane-associated guanylate kinase (MAGUK) family proteins synapse-associated protein 97 (SAP97) and post synaptic density protein 95 (PSD-95), and also recruits protein kinase A (PKA), protein kinase C (PKC), and calcineurin (CaN) in appropriate synaptic domains to regulate excitatory synaptic strength [2-9].
AKAP79/150 also coordinates signal transduction at different subcellular locations and participate in regulating normal physiological functions of cells [10]. AKAP79/150 organizes Ca2+/calmodulin-dependent protein phosphatase (PP2B, also known as CaN), calmodulin, cAMP-dependent PKA, PKC, and the transcription factor nuclear factor of activated T cells (NFAT) into a membrane-delimited signalosome at the plasma membrane [11-13]. Upon Ca2+ store depletion, the stromal-interactingmolecule-1 protein on the plasma membrane targets Orai1 to AKAP79 signaling complex, which interacts with the AKAR on the N-terminus of the Orai1 Ca2+ channel [14-16]. The Ca2+ spikes can transmit signals from the distal dendrite to the nucleus by stimulating CaN to dephosphorylation and activating NFAT [12, 17-23]. The neuronal activity regulated by M current induces NFAT-mediated transcriptional up-regulation of KCNQ channels and inhibits neuronal excitability. This signal pathway limits neuronal hyperexcitation through negative-feedback, which is the basis of myriad diseases such as chronic pains, epilepsy and cardiovascular dysfunction [24].
Accumulating evidence suggests that aberrant expression of AKAP79/150 contributes to the pathophysiological mechanisms of various human disorders. Both chronic restraint stress and unpredictable chronic mild stress can increase the expression level of AKAP150 protein in the basolateral amygdala of depressive mice and induce its redistribution into the synapses. Moreover, studies have also shown that the expression of AKAP150 in dorsal root ganglia neurons of inflammatory pain mice is significantly increased, and the pain response is reduced in Akap5 knockout mice [25]. In addition, the palmitoylation of AKAP150 is significantly increased in the hippocampus of epileptic mice [26], and inhibition of palmitoylation of AKAP79/150 is helpful to the anticonvulsive effect of valproic acid, a seizure treatment drug [27].
In this review, we present an overview on the physiological roles of AKAP79/150 in the brain and deregulations of AKAP79/150 expression and its substrates in various disorders, like inflammatory pain, epilepsy, and depression, providing a new entry point for the diagnosis and treatment intervention for these disorders.
2. Biological Characteristics of AKAP79/150
2.1. Overview of AKAPs
AKAPs belong to a family of scaffolding proteins, which anchor PKA and other important proteins, including protein kinase, protein phosphatase, phosphodiesterase, G protein coupled receptors (GPCRs) [10, 28]. AKAPs are also a group of functionally related regulatory proteins with different structures, which play an important role in ensuring the accuracy of intracellular PKA-dependent signaling pathways and coordinating the precision of signal transduction at different subcellular [29]. Therefore, AKAPs are not only widely involved in the regulation of normal physiological functions of cells, but also play a vital role in various human diseases [10].
Indeed, more than 70 different AKAPs have been isolated and identified so far [10]. AKAPs can form multi-protein complexes in different subcellular regions that integrate the cyclic adenosine monophosphate (cAMP) signaling with pathways. Evidence suggests that AKAP can recognize and specifically anchor activated GPCR. Subsequently, other proteins anchored to AKAP are sequentially activated to generate, utilize, degrade, and regulate the synthesis of cAMP [30]. In addition, cAMP activation of PKA anchoring to AKAPs is crucial for regulating human dendritic cells lipid rafts antigen presentation [31, 32]. AKAPs signaling complexes have been identified as crucial regulators of a variety of glutamate receptors and ion channels [8, 33, 34]. In the past decades, as a member of the AKAPs, many studies have focused on the distribution and function of AKAP79/150 in the brain [8, 35, 36].
2.2. Structure of AKAP79/150
AKAP79/150 is a prominent synapse-targeted AKAP, which is mainly expressed in the central nervous system and play an important role in signal transmission and synaptic plasticity [37]. AKAP150 is anchored by binding the hydrophobic surface of an amphiphilic helical molecule in its molecule to the N-terminal of the PKA regulatory subunit dimer (RII) [38]. Furthermore, in addition to targeting PKA, AKAP150 also has binding sites with other signal molecules, such as PKC, PP2B, some membrane receptors, and ion channels, so that different signal pathways can interact with each other, which is conducive to the integration and transmission of specific information in cells [39, 40].
AKAP79/150 contains a unique targeting sequence near the N-terminal, also known as the targeting domain. In this domain, AKAP79/150 has three different basic targeting sub-domains (A, B, and C domains) (Figure 1). Three membrane-bound basic regions can bind with phosphatidylinositol 4,5-bisphosphate (PIP2) and target specific subcellular regions, making PKA very close to its specific substrate molecules, and the catalytic reaction can be carried out efficiently [40, 41]. AKAP79/150 binds to PKC through the A sub-domain of the basic N-terminal targeting domain [42], but binds to a variety of adenylyl cyclases (ACs) isomers through the B sub-domain of the targeted domain [43, 44].
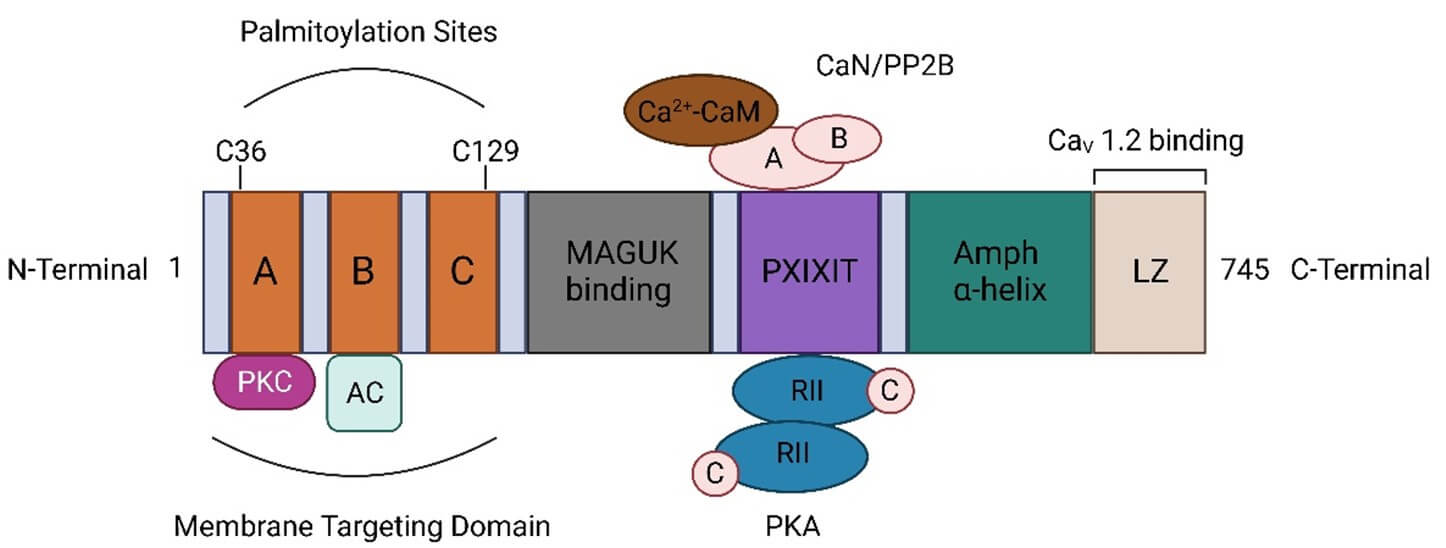
AKAP79 anchors CaN through the 'PxIxIT' type docking motif near the C-terminal [41, 45, 46]. Between the targeting domain and the CaN anchoring domain, AKAP150 has an internal structure that binds to MAGUK. In addition, through the PDZ (an initialism combining the first letters of the first three proteins discovered to share the domain-PSD-95, Drosophila disc large tumor suppressor (DLG1), and Zonula occludens-1 protein (ZO-1)) domains in MAGUKs, AKAP150 is connected to the NMDARs and the C-terminal of AMPARs [47-50].
2.3. Function of AKAP79/150
AKAP79/150 can combine with GPCR superfamily members β2-adrenergic receptor [51, 52], orchestrating the interaction of F-actin [53], PIP2 [41], MAGUKs [48], cadherin cell adhesion molecule [54, 55], ACs [43, 56, 57], protein kinase [1], and protein phosphatase [58], and regulating active-dependent synaptic transmission by connecting with AMPA receptors [48], heat-activated transient receptor potential family V type 1 (TRPV1, also known as the capsaicin receptor) channels [59], L-type calcium (CaV) channels [60], M-type potassium (KCNQ, Kv7) channels [61], and acid sensing ion channels [62] in PSD. PSD scaffolding proteins are an important structural basis of synaptic transmission and play an important supporting role in the integration of multiple receptors and signal molecules [63]. A variety of protein kinases and protein phosphatases, including PKA, PKC, and CaN, are integrated into the PSD region via AKAP150 and play an important regulatory role in the dendritic spine stability [49].
cAMP-dependent PKA is a tetrameric molecule consisting of a tetramer that contains a regulatory (R) subunit dimer, which include four different phenotypes of RIα, RIβ, RIIα and RIIβ [64-68], and two catalytic (C) subunits [69]. When the body is stimulated by the outside world, it will trigger a signal cascade reaction and activate the cAMP signaling pathway. The activated cAMP is combined with the R subunit of PKA, causing the C subunit to be released from PKA and activated. The activated catalytic subunit plays a role in signal transduction. PKA-mediated signaling pathways play an important role in the function of many neurons and the formation of learning and memory, and are also involved in various forms of synaptic plasticity [2, 70].
The PKC family of phospholipid-dependent serine/threonine kinase consists of different isozymes (PKCα, PKCβI, PKCβII, PKCγ, PKCδ, PKCε, PKCη, PKCθ, PKCζ, and PKCι/λ) [71, 72], all of which have a R domain that can bind to PKC activators and a C domain, and variable regions [72]. The function of PKC is to phosphorylate other proteins by interacting with them to participate in synaptic plasticity [73, 74]. PKC also modulates the targeting of the NR1 subunit of NMDAR to the postsynaptic membrane [75]. The serine/threonine phosphatase CaN holoenzyme is a heterodimeric protein, consisting of a C subunit A and a R subunit B [76, 77]. CaN is not only important for cardiac development, pathophysiology, and nervous system development [78], but also as a structural protein to control synaptic function and behavioral learning [79]. Interestingly, a recent study discover that CaN can directly suppress the activity of PKA when removing phosphate from substrates initiated by PKA, thereby avoiding the costly and persistent ineffective cycle of phosphorylation and dephosphorylation of PKA and CaN at these sites [80].
AKAP79/150 can regulate the phosphorylation and dephosphorylation of various ionotropic receptors by anchoring PKA, PKC, and CaN in the postsynaptic membrane [24, 39, 43, 81, 82], thereby affecting the activity and transport of ion channels. In the basal state, AKAP79/150 bound to a pool of largely inactive CaN in synapses. During NMDAR-dependent long-term depression (LTD), AKAP79/150 binds to PSD-95, causing the release of CaN, which helps to NMDAR-triggered an enhancement of synaptic AMPAR endocytosis [83, 84]. In addition, AKAP79/150-anchored PKA and CaN can control the recruitment or removal of Ca2+-permeable AMPA-type glutamate receptors at hippocampus synapses by regulating GluA1 serine845 (S845) phosphorylation during long-term potentiation (LTP) / LTD, playing key antagonistic roles [83, 85-89].
Indeed, L-type Ca2+ channels and AMPA/kainate receptors are modulated by KA [35, 90, 91], while NMDARs are phosphorylated by PKC [92, 93] and dephosphorylated by CaN [94-96]. PKA-mediated phosphorylation of GluA1 serine 845 (S845) has been shown to promote GluA1 cell-surface targeting and synaptic retention, increase channel open-probability, and facilitate the induction of LTP through AKAP5 [97-103], while dephosphorylation of GluA1 S845 is associated with endocytosis and LTD [98, 100, 102, 104-107]. CaMKII/PKC-mediated phosphorylation of GluA1 serine 831 (S831) increases channel conductance and regulates LTP [100, 104, 108-111]. Finally, PKC-mediated phosphorylation of GluA2 serine 880 (S880) disrupts the interaction between GluA2 and GRIP, allowing for AMPAR endocytosis and LTD [47, 112, 113].
2.4. Distribution of AKAP79/150
AKAP79/150 is widely distributed in the nervous system. In the peripheral nervous system, AKAP150 is mainly expressed in sensory neurons of dorsal root ganglion (DRG) [25, 114], and locates in the plasma membrane of the soma, axon initiation segment, and small fibers. Most of these neurons are nociceptive afferent C fibers and a small portion is Aδ-fibers [115]. In the central nervous system, AKAP5 is detected in an increasing number of tissues and cells. AKAP5 is widely distributed in the brain [116-122], heart [123], arterial smooth muscle [124, 125], pancreas [122], liver [116, 122], skeletal muscle [122], uterus [126], stomach [127], parotid [128, 129], diploid fibroblasts [130], amnion fibroblasts [131], T cells [132], red nucleus [133], Purkinje cells, olfactory bulb neurons, basal ganglia, and cortical actin cells [134]. Intriguingly, studies have found striking differences in AKAP150 between brain regions in mouse [135]. Among them, striatum and cerebral cortex had the highest expression levels, followed by hippocampus and olfactory bulb, while cerebellum, hypothalamus, and brain stem had low expression levels [135].
2.5. Post-Translational Modification of AKAP79/150
Before proteins become biologically active, they undergo a series of complex modification processes such as phosphorylation, glycosylation, ubiquitination, and lipidation. Protein palmitoylation is the most common and only reversible post-translational lipid modification, usually refers to the covalent binding of 16-carbon palmitic acid to the side chain of protein specific cysteine residues (Cys) via a labile thioester bond, which is of great significance for protein trafficking, localization in cells and function [136-138].
Protein palmitoylation is achieved by palmitoyl transfer mediated by protein acyltransferase, and its activity is mainly mediated by the structure of DHHC (Asp-His-His-Cys) [139, 140]. These enzymes are distributed in different tissues, mainly located in Golgi apparatus, endoplasmic reticulum or plasma membrane [141]. Currently, 23 DHHC enzymes have been reported, which play an important role in neuronal development and synaptic plasticity [142].
Of note, ZDHHC2 is a member of the important DHHC family in neuroendocrine cells [143], which can mediate palmitoylation of PSD-95 and AKAP79/150, impacting synaptic targeting of AMPARs [144, 145]. Studies have shown that ZDHHC2 can catalyze the S-palmitoylation of two conserved cysteine residues (Cys36 and Cys129) within the A and C basic regions of AKAP79/150 [146]. Although this modification is not important for the localization of AKAP79/150 in the plasma membrane and the connection with F-actin, it is required for the localization of AKAP79/150 to the recycling endosomes, and is closely associated with its localization to cholesterol rich, anti-detergents lipid rafts [141, 147, 148]. Additionally, it is worth noting that PSD is an important structure for anti-detergents and therefore palmitoylation of many PSD proteins is closely related to their synaptic localization [50].
One recent study showed that the palmitoylation of AKAP79/150 modulates its postsynaptic nanoscale organization, trafficking, and mobility in hippocampal neurons [149]. In cultured hippocampal neurons, researchers have observed that the palmitoylation of AKAP79/150 is bidirectionally regulated by synaptic activity, thereby coordinating receptor exocytosis, synaptic spine morphological changes, GluA1 membrane surface expression and AMPAR synaptic activity closely related to LTP and LTD [50, 149]. The literature reported that CP-AMPARs containing GluA1 and lacking GluA2 play an important role in the formation of LTP. Palmitoylation of AKAP79/150 not only regulates synaptic transmission of CP-AMPARs under normal conditions, but also is critical for Ca2+-permeable AMPA-type glutamate receptors -dependent LTP [26, 50, 150].
On the other hand, protein depalmitoylation is the process by which palmitic acid is removed from modified proteins and contributes. Similarly, protein depalmitoylation also serves as particularly crucial regulators of protein function in neurons. The research shows that CaMKII-mediated autonomous phosphorylation and depalmitoylation are required for the synaptic removal of AKAP79/150 accompanied by the shrinkage of the volume of dendritic spines after the induction of LTD [151, 152]. In addition, synaptic removal of AKAP79/150 can prevent PKA-mediated re-phosphorylation of AMPAR, promoting endocytosis of AMPAR, which weakens the strength of the synapse [151]. Of note, the fully phosphorylated AKAP79 mutant showed a significant decrease in palmitoylation levels compared with the partially phosphorylated mutations [151].
3. Potential Role of Akap79/150 in Human Disorders
So far, many studies have shown that the dysfunction of AKAP79/150 expression plays an important regulatory role in a variety of neural physiological activities, especially in some major neuropsychiatric diseases.
3.1. Inflammatory Pain
3.1.1. Inflammatory Pain Overview
Inflammatory pain is a severe chronic pain that is caused by inflammation caused by trauma, bacterial and viral infections, and surgical procedures. Allodynia, hyperalgesia and sensitization are its main clinical manifestations [153]. Injury or inflammation leads to the release of inflammatory mediators such as bradykinin, prostaglandin E2 (PGE2), L-glutamate and nerve growth factor (NGF), which activate “pain-” sensing neurons, increase nociceptor responsiveness and lower the threshold for pain [154-157].
Inflammatory pain not only severely affects the quality of life, but also increases the incidence of mental diseases [158]. Therefore, it is of great significance to further study its pathogenesis and develop corresponding analgesic drugs in clinical treatment. Of note, some important discoveries in this field in recent years are closely related to the AKAP79/150.
3.1.2. Potential Mechanism of AKAP79/150 in Inflammatory Pain
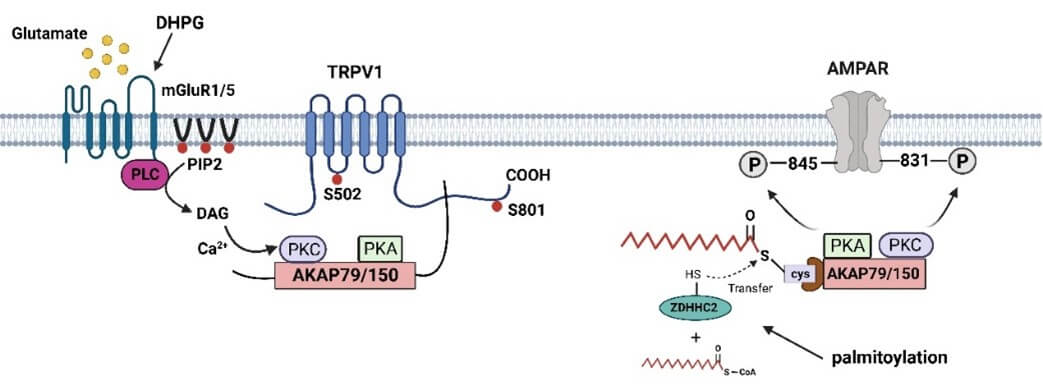
The nociceptive transduction molecule TRPV1 is the best-studied TRP channel, which plays a crucial role in hyperalgesia [159]. Activation of PKA or PKCε by inflammatory mediators phosphorylates TRPV1 and increases the sensitivity of channel gating, depending not only on the ability of the associated these kinases to bind AKAP79/150, but also on the ability of AKAP79/150 to bind TRPV1 [59, 160].
Previous studies have demonstrated that AKAP150-anchored PKA and PKC control the phosphorylation and functional status of TRPV1 [161, 162], but the scaffolding protein is not involved in PP2B-driven channel desensitization [76]. Although both AKAP150 and the PKA regulatory subunit PKA RIIα are associated with TRPV1, the association between PKA and TRPV1 is mediated by AKAP150. Disruption of PKA binding to AKAP150 strongly attenuated the reduction of TRPV1 desensitization and hyperalgesia induced by PGE2 [163].
Blocking binding or removing AKAP79/150 inhibits the sensitization of TRPV1 [59]. The siRNA against AKAP150 effectively blocked the NMDA-induced phosphorylation of S800 of TRPV1 in trigeminal ganglia [164]. In addition, phosphorylation of the S502 site on TRPV1 as the main mechanism by which AKAP79 mediates translocation of TRPV1 to the membrane [59]. A 14 aa domain, corresponding to residues 736–749 mediating AKAP79 binding was found in the C-terminal domain of TRPV1 [59]. In fact, critical residues within this binding site, namely D738, R740, C742, and V745 have been identified [165]. If binding is blocked, the sensitization effect was abrogated. 736–745-TAT (a membrane-permeable decoy peptide) attenuated the thermal hyperalgesia induced by Formalin was attenuated, as well as the mechanical hyperalgesia induced by Group I metabotropic glutamate receptors (mGluR1/5) agonist (DHPG) [165, 166]. In turn, amino acids 326–336 on AKAP79 responsible for its interaction with TRPV1. the binding site of TRPV1 on AKAP79 a sequence of 11 aa, 326–336 in the C-terminal. The sensitization of TRPV1 by PKC and PKA in vitro and inflammatory hyperalgesia in vivo were reduced by using a peptide mimicking the 326–336 site on AKAP79 [167]. Thus, antagonizing the interaction between AKAP79 and TRPV1 may offer a promising analgesic strategy.
As mentioned earlier, L-glutamate mediate hyperalgesia. mGlu1/5 couple activation of phospholipase C (PLC), resulting in release of AKAP150 from cell membrane of the PIP2-rich region can associate with target substrate proteins, including TRPV1 [168]. There was a twofold increase in AKAP association with TRPV1 following DHPG treatment [169]. AKAP5 KO mice with inflammation did not facilitate DHPG-induced pain behavior and the proportion of S800-phosphorylated TRPV1 was significantly reduced in DRG neurons from AKAP5 KO mice after NGF treatment [25]. Moreover, a functional transcriptional link has been observed between serum response factor (SRF) and AKAP150. DHPG-stimulated upregulation of AKAP150 was blocked pretreated with SRF siRNA ([170].
AKAP150 trafficking and synaptic localization is modulated by palmitoylation [26]. A recent study showed that AKAP150 translocated from cytoplasmic to synaptic site after intraplantar injection of complete freund’s adjuvant (CFA). The palmitoylation levels of AKAP150 were significantly increased, and mediated the accumulation and phosphorylation of GluA1. Inhibition of AKAP150 palmitoylation through intrathecal injection palmitoylation-deficient AKAP150 mutant vectors, AKAP150 (C36, 123S) or ZDHHC 2-siRNA or 2-bromopalmitate (2-BP) could relieved inflammatory pain [171].
Noxious thermal, mechanical, or chemical stimuli lead to depolarization of nociceptive sensory neurons, triggering a series of events such as calcium influx, Ca2+/CaM–dependent activation of ACs, and intracellular cAMP production [172-174]. AKAP79/150-anchored AC is vital for the sensitization of TRPV1 to cAMP/PKA-dependent regulation [175]. Furthermore, AC5/6 interaction with AKAP150 is necessary for persistent nociceptor spontaneous activity [114]. Genetic deletion of AC1 prevented the translocation of AKAP79/150 and PKA, as well as upregulation of synaptic GluA1-containing AMPARs in the insular cortex after nerve injury [176].
AKAP79/150 anchoring of kinases and phosphatase control ion channels response to activation signals [177]. There are 63.0%, 57.6%, and 11.8% of APAP150-positive neurons were co-expressed with CaV1.2, voltage gated sodium channel, and Kv1.2 in rat DRG neurons, respectively. Studies in small nociceptive DRG neurons demonstrated that AKAP150 interacts with TRPV1 and CaV1.2 in the soma and axon initial segment [115]. Depolarization of sensory neurons rapidly activates PKA type II (PKA-II) in nociceptors by Ca2+ influx through CaV1.2 channels [178]. BAPTA-AM-driven chelation of intracellular Ca2+ not only increased PKA-mediated sensitization of TRPV1, but also increase the association of AKAP150 with TRPV1 [179]. CaM serves as a part of a calcium-sensitive complex was able to bind to C terminus 35-aa segment of TRPV1 and effectively prohibits AKAP150 from associating with TRPV1 [179, 180]. Another low voltage-gated (T-type) Ca2+ channels isoform, CaV3.2 was found to be able to immunoprecipitated with AKAP150. The increased phosphorylation of Ser/Thr residues in CaV3.2 induced by dibutyryl cAMP can be reversed by AKAP St-Ht31 inhibitory peptide ([181].
Like TRPV1, TRPV4 was shown an interaction with AKAP79 by co-immunoprecipitation in small nociceptive sensory neurons [59]. Previous study has demonstrated that AKAP79 plays a critical role in tethering PKA and PKC to TRPV4 to modulate its gating [182]. Modulating TRPV4 function using peptide antagonists targeting the interaction between TRPV4 and AKAP79 is considered to be an effective therapeutic strategy [183].
AKAP150 is also involved in chemotherapy-induced neuropathic pain (CINP). Knocked down AKAP150 by intrathecal injection of AKAP150 siRNA or AAV5-Cre-GFP virus significantly alleviated paclitaxel-induced hypersensitivity, and partially restored CaN phosphatase activity and IL-4 expression [184]. In addition, AKAP150 mediates the sensitization of transient receptor potential anchor protein 1 (TRPA1) regulated carboplatin-induced mechanical allodynia and cold hyperalgesia [185]. These findings indicate that AKAP150 plays an important role in the neuropathic pain (Figure 2).
3.2. Epilepsy
3.2.1. Epilepsy Overview
Epilepsy is a common chronic neurological disease characterized by epilepsy, which is caused by overexcitation and synchronous abnormal discharge of brain neurons. The formation of neuronal network is closely related to synaptic transmission. When the synaptic transmission function is abnormal, the equilibrium between excitation and inhibition of the neural network is destroyed, which induces abnormal firing of neurons and finally leads to seizures [186]. Despite more than 40 kinds of antiseizure drugs have been used in clinic in recent years, about one-third of patients are still unable to effectively control epileptic seizures [187].
The etiology of epilepsy is complex and diverse, involving structural, genetic, metabolic, infectious, or immune factors [188]. The abnormal firing of neurons is found to be closely related to dysfunction of the mitochondrion and abnormalities of neurotransmitters and ion channels [189-191]. When neurons in the brain discharge abnormally, motor, sensory, cognitive, psychic and autonomic nervous functions will be impaired [192]. Intriguingly, studies have found that AKAP79/150 may play an important role in the onset and development of epilepsy.
3.2.2. Potential Mechanisms of AKAP79/150 in the Development of Epilepsy
It was previously shown that AKAP150 knockout mice show learning disabilities and are resistant to seizure after peritoneal injection of pilocarpine (300 mg/kg) [193]. In particular, kainic acid (KA)-induced seizures have been demonstrated to increase palmitoylation of AKAP150, thereby promoting movement to postsynaptic lipid rafts [26]. Hence, AKAP150 may have a significant role in the pathogenesis of epilepsy.
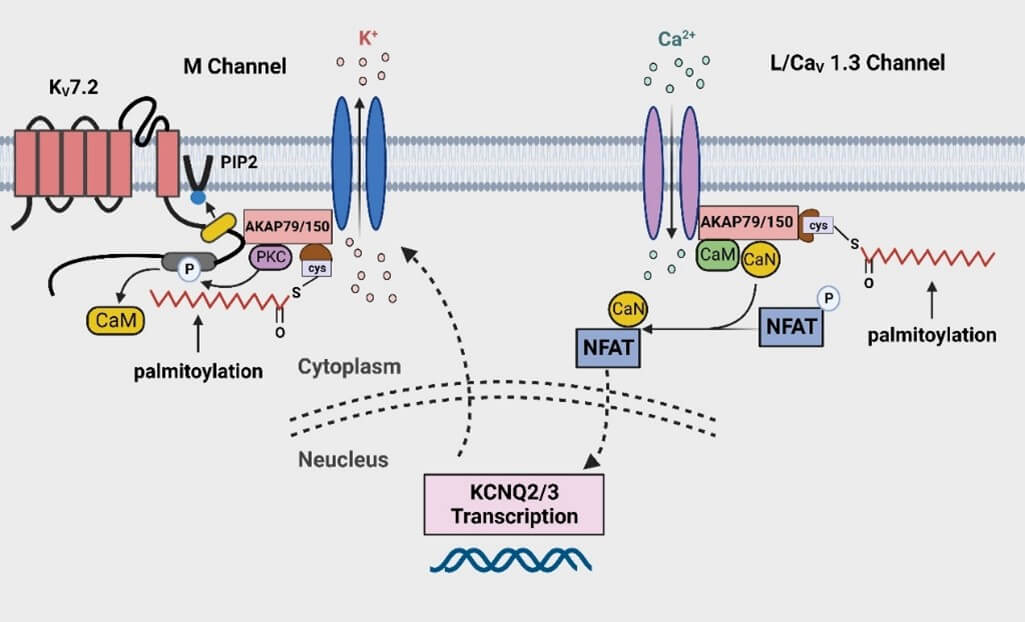
On the one hand, the KCNQ2/3 mutation, which leads to a mild decrease of M-channel activity, is associated with the onset of benign neonatal epilepsy. These mutations can reduce the excessive excitability responsible for epileptic seizures [194]. The palmitoylation of AKAP79/150 has been proved to be critical to its mediated regulation of KCNQ2 both in vivo and in vitro experiments, and M-current suppression is involved in the pathophysiology of seizures [27]. Seizures activate Gq-coupled receptors, and Kv7.2 subunit is phosphorylated by AKAP79/150-anchored PKC. Phosphorylated Kv7.2 subunits release calmodulin (CaM), thereby reducing the binding of PIP2 to Kv7.2 subunit. Suppressed M-current reduces channel functionality, ultimately leading to neuronal hyperexcitability [193, 195-199]. Due to the palmitoylation modification of AKAP79/150 being necessary for phosphorylation of Kv7.2 subunit by AKAP79/150-anchored PKC, the anticonvulsant effect of valproic acid is achieved by interfering with the palmitoylation of AKAP79/150 to prevent the M-current from suppressive neurotransmitters during seizures, thereby preventing the progression of seizures [27, 199].
On the other hand, AKAP79/150 regulates the transcriptional expression of KCNQ2/3 by coordinating L-type voltage-gated Ca2+ channels to limit the overexcitation of the nervous system [24]. After induction of seizures by pilocarpine and KA, the opening of CaV1.3 channels produces an elevated local Ca2+i signals, activating AKAP79/150-anchored CaN associated with CaV1.3 channels. NFAT is then dephosphorylated and activated by Ca2+-CaM/CaN, causing NFAT to translocate from the cytoplasm to the nucleus, acting on the KCNQ2/3 gene regulatory elements and upregulating M-current. The increased expression of KCNQ2/3 channels serves as a negative-feedback loop to suppresses the increase in neuronal excitability [24,200].
The above findings suggest a crucial role of AKAP79/150 and its palmitoylation in the pathophysiology of epilepsy (Figure 3). However, extensive research is needed to clarify the potential molecular mechanisms of AKAP79/150 in the development of epilepsy.
3.3. Depression
3.3.1. Depression Overview
Depression is a common mental disorder characterized by persistent low mood, intellectual disability, cognitive impairment, volitional decline, and impaired social function, affecting more than 300 million worldwide [201, 202]. The etiological hypotheses of depression include the monoamine neurotransmitter hypothesis, the hypothalamic pituitary adrenal axis dysfunction hypothesis, the neural plasticity and neurotrophic factor hypothesis, the inflammation and cytokine hypothesis, and the intestinal flora imbalance hypothesis [203-208]. In the past decades, however, the pathogenesis of depression still remains poorly understood, and currently there is no cure for depression. Impressively, the role of AKAP79/150 in the development of depression advances our understanding of this disease.
3.3.2 Potential Mechanisms of AKAP79/150 in Depression
The synaptic dysfunction has been extensively studied in patients with depression and in animal models [209-211]. The glutamate-mediated excessive activation of extra synaptic NMDAR has been found to be closely related to animal behaviors such as decreased food intake, weight loss, loss of pleasure, cognitive impairment, and social disorder, which are also common clinical manifestations of patients with depression [212-216].
A study identified AKAP5 copy-number increases using DNA copy-number analysis from the human brain DNA samples of post-mortem bipolar disorder and schizophrenia patients [217, 218]. Of interest, the numerical density of AKAP5-expressing neurons was significantly increased in the left and right anterior cingulate cortex of patients with bipolar disorder [219]. The DNA copy variants in eight AKAP5 were also found in individuals with autism spectrum disorders [220].
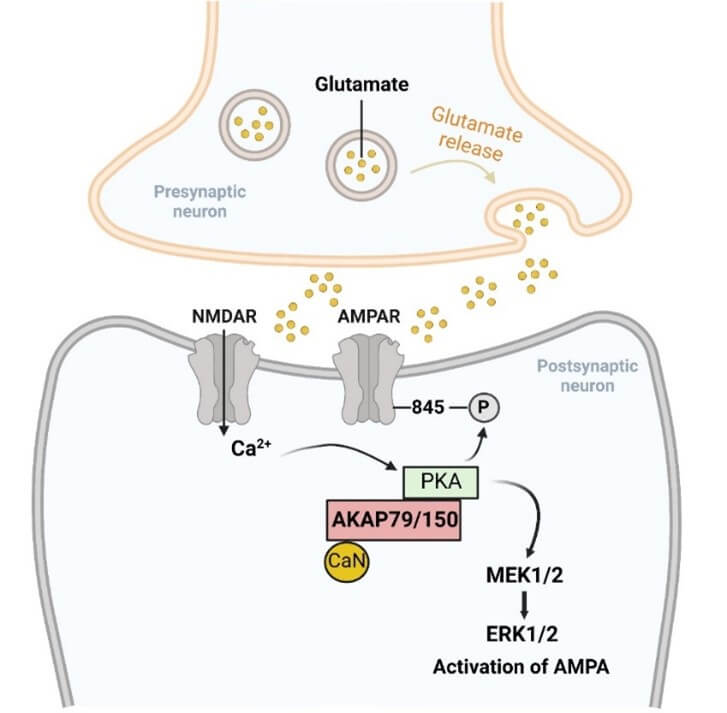
Another study found that aberrant expression of Akap5 is found in amygdala after unpredictable chronic mild stress-induced depression-like behavior in rats, and antidepressants could reverse the expression of Akap5 [221]. Furthermore, a recent study shows that both chronic restraint stress and unpredictable chronic mild stress increase the expression of AKAP150 and induce its redistribution into the synapses in the basolateral amygdala of depressive mice [222]. As a AKAP79/150 anchoring kinase in SH-SY5Y neurons, PKA is recruited to the synaptic compartment, facilitating the phosphorylation of GluA1 Ser845 site and the insertion of GluA1-containing AMPARs into the neuronal postsynaptic membrane [49, 222, 223]. AMPAR transported to the postsynaptic membrane is located on the PSD-95 scaffold and mediated glutamatergic synaptic transmission contributes to depressive-like behaviors [222, 224]. Interestingly, studies have found that curcumin, a multitarget drug with antidepressant effect, can protect neurons from glutamate insult by reducing Ca2+ influx induced by NMDAR and blocking the translocation of AKAP79 from cytomembrane to cytoplasm. At the same time, curcumin facilitates the phosphorylation of AMPAR and its downstream signal transmission from MEK1/2 to ERK1/2 in PKA dependent manner [223]. These findings reveal insight into the pathophysiology of depression and provide a novel target for the development of antidepressants (Figure 4).
3.4 Diabetes
Interestingly, more recently has found that the adipocyte hormone leptin could not promote KATP channel trafficking and membrane hyperpolarization in human β-cells from obese type II diabetic donors and β-cells from obese diabetic db/db mice lacking functional leptin receptors; however, leptin activates Src kinase, and AKAP79/150 anchors and activates PKA after Src kinase activation, initiating NMDAR-CaMKKβ-AMPK signaling cascades and reactivates the action of leptin. PKA-dependent actin remodeling promotes KATP channel trafficking and increase in K+ conductance, causing β-Cells hyperpolarization and inhibit glucose stimulated insulin secretion [37]. Noteworthy, studies have found that insulin secretion can be regulated by the reversible phosphorylation of β cell proteins through the AKAP79/150 targeting of PKA and PP2B [37, 122, 225]. In addition, studies have found that separately selective down-regulation of KV2.1 and large conductance Ca2+-activated K+ channel by activating AKAP150-CaN dependent NFATc3 signaling and the β1 subunit of the large conductance Ca2+-activated K+ channel in vascular smooth muscle cells can enhance arterial tone during diabetes [226-228].
In another study of diabetic hyperglycemia, it was found that when the extracellular glucose level increased, P2Y11 receptor activated, promoting AC5 activity and local cAMP production. This cAMP microdomain can enable AKAP5 to recruit PKA that is intimately associated with L-type CaV1.2 channels to increase their phosphorylation at serine 1928 (S1928), which will potentiate channel activity [229, 230]. Intriguingly, the selective P2Y11 agonist NF546 increases cAMP levels and CaV1.2 channel activity in human and mouse vascular smooth muscle cells, thus exerting a similar effect as increasing glucose. Counterintuitively, genetic ablation of AKAP5 (AKAP5-/-) inhibited the increase in cAMP and CaV1.2 channel activity induced by elevated glucose and the NF546. In addition, AKAP5-/- can completely abrogated the association of P2Y11 receptors with PKA and the CaV1.2 α1C subunit, as well as the CaV1.2 α1C subunit with AC5 and PKA with AC5 in vascular smooth muscle cells. Altogether, the results suggested the AKAP5/P2Y11/AC5/PKA/CaV1.2 signaling complex may be targeted for the treat diabetic vascular complications [231, 232].
3.5. Cardiovascular Disorders
AKAP79/150 plays an important role in heart function mainly by regulating the calcium ion changes in the heart, myocardial hypertrophy, and heart failure. On one hand, previous study found that Akap5-knockout mice have significant cardiac hypertrophy, and carvedilol can reverse cardiac hypertrophy and cardiac insufficiency in Akap5 gene knockout mice by regulating the activity of CaN and CaMKII [28, 233]. It is worth noting that recent studies have found that Selective β1-adrenergic receptor blocker metoprolol reduces ischemic cardiac remodeling and fibrosis by improving cardiac AKAP5 expression and AKAP5-PP2B interaction [234]. Studies have shown that chronic activation of CaN is associated with cardiac hypertrophy and a secondary enhancement of intracellular Ca2+ treatment that is tied to the hypertrophy response itself, while targeted inhibition of CaN alleviates cardiac hypertrophy in vivo [235-239]. In addition, AKAP79/150 regulates β-adrenergic receptor signaling, trafficking, and recycling by anchoring to PKA or CaN exert cardioprotective effects [240, 241]. Interestingly, a recent study suggests that AKAP5 may anchor CaN to regulate NFATc3 remodeling in H9c2 cardiomyocytes exposed to hypoxia and reoxygenation after ischemia-reperfusion injury [242]. Furthermore, AKAP150-anchored CaN acute activation transcription factor NFATc3 mediates voltage gated K+ currents downregulation in ventricular myocytes following myocardial infarction, increasing the probability of arrhythmias [243]. On the other hand, by anchoring and activating conventional PKCs, AKAP150 promotes the activation of NF-κB, and mediates the toxic cardiac effects of hyperglycemia [244, 245].
3.6. Alzheimer’s Disease
Previous studies have shown that AKAP79 is highly expressed in cortical regions and hippocampal subregions that are susceptible to the development of neurofibrillary pathology in alzheimer’s disease [42, 117, 246]. In addition, the activity of CaN, which binds and localizes with AKAP79, decreases in alzheimer’s disease [42, 246]. Interestingly, dysregulation of subcellular localization of PKA-Cβ, PKA-RIIβ, and AKAP79 exist in alzheimer’s disease, which may allow for the specific targeting of tau protein by activated PKA after elevations in cAMP levels. Hyper-phosphorylation of the tau protein can form neurofibrillary tangles in the brain of alzheimer’s disease patients [247, 248]. It has been reported that AKAP150 may coordinates PKA and CaN regulation of CP-AMPARs to mediate disruption of hippocampal neuronal plasticity and LTP/LTD balance by Aβ oligomers in Alzheimer’s disease, thus impairing learning and memory [150, 249].
3.7. Hypertension
Research has found that when the angiotensin II signal is elevated, activation activates Gq coupled receptors, increasing cytosolic DAG and IP3 levels. DAG activates PKC and CaN anchored by AKAP150. When PKC is activated, it can phosphorylate nearby TRPV4 and CaV1.2 channels, increasing the probability of their opening. The opening of the TRPV4 channel develop stuttering persistent Ca2+ sparklets signals in arterial myocytes that regulate Ca2+ influx and NFATc3-dependent gene expression in smooth muscle. This increases arterial [Ca2+]i and myogenic tension, ultimately leading to hypertension [124, 125, 250-255].
3.8. Cancer
Recently, an analysis using chi-square and Fisher exact test found that AKAP5 expression was decreased in non-mucin producing stomach adenocarcinoma based on the cancer genome atlas data [256]. In addition, using GSEA to analyze the cancer genome atlas dataset, it was found that gene sets related to cholesterol homeostasis, glycolysis, estrogen response late, adipogenesis, estrogen response early, notch signaling, and peroxisome were differentially enriched with the low AKAP5 expression phenotype. It suggested that these may be the key pathways regulated by AKAP5 in non-mucin producing stomach adenocarcinoma [256].
Moreover, it was found that the nonsynonymous coding mutations of AKAP5 was enriched in the metastatic tumor of primary breast cancers with paired metastatic lesions by exome sequencing. Interestingly, further exploration using the cancer genome atlas dataset found that AKAP5 was expressed at lower levels in the subtypes (basal-like and HER2-enriched) of breast cancer with highest risk of recurrence. Accordingly, low expression of AKAP5 is more likely to recurrence and metastasis in breast cancer [257].
4. Conclusion
A growing body of evidence has suggested the crucial role of AKAP79/150 in the pathophysiological conditions of some human disorders. Notably, increased AKAP150 expression is closely associated with the pathogenesis of depression. Pharmacological targeting of AKAP150 expression has shown great promise for the depression treatment. In addition, alterations of AKAP150 expression also exist in patients with epilepsy or epileptic animal models, neuropathic pain, or schizophrenia. These lines of evidence highlight the important roles of AKAP79/150 in the progression of these disorders, and render AKAP79/150 as a valuable therapeutic target. Together, the contributions of the AKAP79/150 in regulating disruptions of the synaptic circuits is critical for our understanding of the pathophysiology of some diseases and attempts for development of novel treatments. We envision that the future advances into the molecular mechanisms of AKAP79/150 deregulation and its involvement in the pathogenesis of those disorders can be a key in developing new promising treatment strategies.
Acknowledgements
This work was supported by the specially appointed medical expert project of the Jiangsu Commission of Health (2019), the Hospital Pharmaceutical Research Project of Jiangsu Pharmaceutical Association and Chia-Tai Tianqing (No. Q202219), the National Natural Science Foundation of China (No. 82174051 and No. 82104316) and the Medical Science and Technique Foundation of Nanjing Health Commission (No. YKK20119).
Authors’ Contributions
Y.-H. H., W.-R. F., and F. C.: conceptualization. C.-C. C. and Y.-H. H.: writing-original draft preparation. C.-Y. W., L. Z., J. X., X.-P. L., and F.C.: writing-review and editing. F. C., Y.-H. H., W.-R. F., L. Z., and X.-P. L.: funding acquisition. All authors have read and agreed to the published version of the manuscript.
Conflicts of Interest
None.
REFERENCES
[1] Naoto Hoshi, Lorene K Langeberg,
Christine M Gould, et al. “Interaction with AKAP79 modifies the cellular
pharmacology of PKC.” Mol Cell, vol. 37, no. 4, pp. 541-550, 2010. View
at: Publisher
Site | PubMed
[2] M Colledge, R A Dean, G K Scott, et
al. “Targeting of PKA to glutamate receptors through a MAGUK-AKAP complex.” Neuron,
vol. 27, no. 1, pp. 107-119, 2000. View at: Publisher Site | PubMed
[3] Steven J Tavalin, Marcie Colledge,
Johannes W Hell, et al. “Regulation of GluR1 by the A-kinase anchoring protein
79 (AKAP79) signaling complex shares properties with long-term depression.” J
Neurosci, vol. 22, no. 8, pp. 3044-3051, 2002. View at: Publisher
Site | PubMed
[4] Seth F Oliveria, Lisa L Gomez, Mark L
Dell'Acqua “Imaging kinase--AKAP79--phosphatase scaffold complexes at the
plasma membrane in living cells using FRET microscopy.” J Cell Biol,
vol. 160, no. 1, pp. 101-112, 2003. View at: Publisher
Site | PubMed
[5] S M Lilly, F J Alvarez, E I Tietz
“Synaptic and subcellular localization of A-kinase anchoring protein 150 in rat
hippocampal CA1 pyramidal cells: Co-localization with excitatory synaptic
markers.” Neuroscience, vol. 134, no. 1, pp. 155-163, 2005. View at: Publisher
Site | PubMed
[6] Mark L Dell'Acqua, Karen E Smith,
Jessica A Gorski, et al. “Regulation of neuronal PKA signaling through AKAP
targeting dynamics.” Eur J Cell Biol, vol. 85, no. 7, pp. 627-633, 2006.
View at: Publisher
Site | PubMed
[7] Valentina Di Biase, Gerald J
Obermair, Zsolt Szabo, et al. “Stable membrane expression of postsynaptic
CaV1.2 calcium channel clusters is independent of interactions with AKAP79/150
and PDZ proteins.” J Neurosci, vol. 28, no. 51, pp. 13845-13855, 2008.
View at: Publisher
Site | PubMed
[8] Jennifer L Sanderson, Mark L
Dell'Acqua “AKAP signaling complexes in regulation of excitatory synaptic
plasticity.” Neuroscientist, vol. 17, no. 3, pp. 321-336, 2011. View at: Publisher
Site | PubMed
[9] Matthieu Dacher, Shawn Gouty, Steven
Dash,, et al. “A-Kinase Anchoring Protein-Calcineurin Signaling in Long-Term
Depression of GABAergic Synapses.” J Neurosci, vol. 33, no. 6, pp.
2650-2660, 2013. View at: Publisher Site | PubMed
[10] Santosh V Suryavanshi, Shweta M
Jadhav, Bradley K McConnell “Polymorphisms/Mutations in A-Kinase Anchoring
Proteins (AKAPs): Role in the Cardiovascular System.” J Cardiovasc Dev Dis,
vol. 5, no. 1, pp. 7, 2018. View at: Publisher Site | PubMed
[11] Neha Patel, Florian Stengel, Ruedi
Aebersold, et al. “Molecular basis of AKAP79 regulation by calmodulin.” Nat
Commun, vol. 8, no. 1, pp. 1681, 2017. View at: Publisher Site | PubMed
[12] Pulak Kar, Yu-Ping Lin, Rajesh
Bhardwaj, et al. “The N terminus of Orai1 couples to the AKAP79 signaling
complex to drive NFAT1 activation by local Ca(2+) entry.” Proc Natl Acad Sci
U S A, vol. 118, no. 19, pp. e2012908118, 2021. View at: Publisher Site | PubMed
[13] Yu-Ping Lin, Erica Scappini, Carlos
Landaverde et al. “Nuanced Interactions between AKAP79 and STIM1 with Orai1
Ca2+ Channels at Endoplasmic Reticulum-Plasma Membrane Junctions Sustain NFAT
Activation.” Mol Cell Biol, vol. 42, no. 11, pp. e0017522, 2022. View
at: Publisher Site | PubMed
[14] Andrew P Braun “Some assembly
required: SOCE and Orai1 channels couple to NFAT transcriptional activity via
calmodulin and calcineurin.” Channels (Austin), vol. 8, no. 5, pp.
383-384, 2014. View at: Publisher
Site | PubMed
[15] Pulak Kar, Krishna Samanta, Holger
Kramer, et al. “Dynamic assembly of a membrane signaling complex enables
selective activation of NFAT by Orai1.” Curr Biol, vol. 24, no. 12, pp.
1361-1368, 2014. View at: Publisher
Site | PubMed
[16] Jill L Thompson, Trevor J
Shuttleworth “Anchoring protein AKAP79-mediated PKA phosphorylation of STIM1
determines selective activation of the ARC channel, a store-independent Orai
channel.” J Physiol, vol. 593, no. 3, pp. 559-572, 2015. View at: Publisher Site | PubMed
[17] Philip J Dittmer, Mark L Dell'Acqua,
William A Sather “Ca2+/calcineurin-dependent inactivation of neuronal L-type
Ca2+ channels requires priming by AKAP-anchored protein kinase A.” Cell Rep,
vol. 7, no. 5, pp. 1410-1416, 2014. View at: Publisher Site | PubMed
[18] Jonathan G Murphy, Jennifer L
Sanderson, Jessica A Gorski, et al. “AKAP-anchored PKA maintains neuronal
L-type calcium channel activity and NFAT transcriptional signaling.” Cell
Rep, vol. 7, no. 5, pp. 1577-1588, 2014. View at: Publisher Site | PubMed
[19] Jonathan G Murphy, Kevin C Crosby,
Philip J Dittmer, et al. “AKAP79/150 recruits the transcription factor NFAT to
regulate signaling to the nucleus by neuronal L-type Ca(2+) channels.” Mol
Biol Cell, vol. 30, no. 14, pp. 1743-1756, 2019. View at: Publisher Site | PubMed
[20] Angela R Wild, Brooke L Sinnen,
Philip J Dittmer, et al. “Synapse-to-Nucleus Communication through NFAT Is
Mediated by L-type Ca(2+) Channel Ca(2+) Spike Propagation to the Soma.” Cell
Rep, vol. 26, no. 13, pp. 3537-3550 e4, 2019. View at: Publisher Site | PubMed
[21] Ga-Yeon Son, Krishna Prasad Subedi,
Hwei Ling Ong, et al. “STIM2 targets Orai1/STIM1 to the AKAP79 signaling
complex and confers coupling of Ca(2+) entry with NFAT1 activation.” Proc
Natl Acad Sci U S A, vol. 117, no. 28, pp. 16638-16648, 2020. View at: Publisher Site | PubMed
[22] Pulak Kar, Pradeep Barak, Anna Zerio.
“AKAP79 Orchestrates a Cyclic AMP Signalosome Adjacent to Orai1 Ca(2+)
Channels.” Function (Oxf), vol. 2, no. 5, pp. zqab036, 2021. View at: Publisher Site | PubMed
[23] Lucile Noyer, Stefan Feske “Straight
from the channel's mouth: AKAP79 links Ca(2+) influx through ORAI1 to NFAT
activation.” Cell Calcium, vol. 99, pp. 102459, 2021. View at: Publisher Site | PubMed
[24] Jie Zhang, Mark S Shapiro
“Activity-dependent transcriptional regulation of M-Type (Kv7) K(+) channels by
AKAP79/150-mediated NFAT actions.” Neuron, vol. 76, no. 6, pp.
1133-1146, 2012. View at: Publisher
Site | PubMed
[25] Takayoshi Masuoka, Yuka Yamashita,
Junko Yoshida, et al. “Sensitization of glutamate receptor-mediated pain
behaviour via nerve growth factor-dependent phosphorylation of transient
receptor potential V1 under inflammatory conditions.” Br J Pharmacol,
vol. 177, no. 18, pp. 4223-4241, 2020. View at: Publisher Site | PubMed
[26] Dove J Keith, Jennifer L Sanderson,
Emily S Gibson, et al. “Palmitoylation of A-kinase anchoring protein 79/150
regulates dendritic endosomal targeting and synaptic plasticity mechanisms.” J
Neurosci, vol. 32, no. 21, pp. 7119-7136, 2012. View at: Publisher Site | PubMed
[27] Hee Yeon Kay, Derek L Greene,
Seungwoo Kang, et al. “M-current preservation contributes to anticonvulsant
effects of valproic acid.” J Clin Invest, vol. 125, no. 10, pp. 3904-3914,
2015. View at: Publisher
Site | PubMed
[28] Xin Li, Shannon M Matta, Ryan D Sullivan,
et al. “Carvedilol reverses cardiac insufficiency in AKAP5 knockout mice by
normalizing the activities of calcineurin and CaMKII.” Cardiovasc Res,
vol. 104, no. 2, pp. 270-279, 2014. View at: Publisher Site | PubMed
[29] Matthew G Gold, Douglas M Fowler,
Christopher K Means, et al. “Engineering A-kinase anchoring protein (AKAP)-selective
regulatory subunits of protein kinase A (PKA) through structure-based phage
selection.” J Biol Chem, vol. 288, no. 24, pp. 17111-17121, 2013. View
at: Publisher Site | PubMed
[30] Raquel Guinzberg, Antonio Díaz-Cruz,
Carlos Acosta-Trujillo, et al. “Newly synthesized cAMP is integrated at a
membrane protein complex signalosome to ensure receptor response specificity.” FEBS
J, vol. 284, no. 2, pp. 258-276, 2017. View at: Publisher Site | PubMed
[31] Robynn V Schillace, Casey L Miller,
Neal Pisenti, et al. “A-kinase anchoring in dendritic cells is required for
antigen presentation.” PLoS One, vol. 4, no. 3, pp. e4807, 2009. View
at: Publisher
Site | PubMed
[32] Robynn V Schillace, Casey L Miller,
Daniel W Carr “AKAPs in lipid rafts are required for optimal antigen
presentation by dendritic cells.” Immunol Cell Biol, vol. 89, no. 5, pp.
650-658, 2011. View at: Publisher
Site | PubMed
[33] C Dart, M L Leyland “Targeting of an
A kinase-anchoring protein, AKAP79, to an inwardly rectifying potassium
channel, Kir2.1.” J Biol Chem, vol. 276, no. 23, pp. 20499-20505, 2001.
View at: Publisher
Site | PubMed
[34] Jie Zhang, Chase M Carver, Frank S
Choveau, et al. “Clustering and Functional Coupling of Diverse Ion Channels and
Signaling Proteins Revealed by Super-resolution STORM Microscopy in Neurons.” Neuron,
vol. 92, no. 2, pp. 461-478, 2016. View at: Publisher Site | PubMed
[35] C Rosenmund, D W Carr, S E Bergeson,
et al. “Anchoring of protein kinase A is required formodulation of AMPA/kainate
receptors on hippocampal neurons.” Nature, vol. 368, no. 6474, pp.
853-856, 1994. View at: Publisher
Site | PubMed
[36] Aixa F Rivera-Pagán, Miguel P
Méndez-González, David E Rivera-Aponte, et al. “A-Kinase-Anchoring Protein
(AKAP150) is expressed in Astrocytes and Upregulated in Response to Ischemia.” Neuroscience,
vol. 384, pp. 54-63, 2018. View at: Publisher
Site | PubMed
[37] Veronica A Cochrane, Zhongying Yang,
Mark L Dell'Acqua, et al. “AKAP79/150 coordinates leptin-induced PKA signaling
to regulate KATP channel trafficking in pancreatic β-cells.” J Biol Chem, vol. 296, pp.
100442, 2021. View at: Publisher
Site | PubMed
[38] M G Newlon, M Roy, D Morikis, et al.
“The molecular basis for protein kinase A anchoring revealed by solution NMR.” Nat
Struct Biol, vol. 6, no. 3, pp. 222-227, 1999. View at: Publisher Site | PubMed
[39] Wei Wong, John D Scott “AKAP
signalling complexes: focal points in space and time.” Nat Rev Mol Cell Biol,
vol. 5, no. 12, pp 959-970, 2004. View at: Publisher Site | PubMed
[40] Matthew G Gold, Florian Stengel,
Patrick J Nygren, et al. “Architecture and dynamics of an A-kinase anchoring
protein 79 (AKAP79) signaling complex.” Proc Natl Acad Sci U S A, vol.
108, no. 16, pp. 6426-6431, 2011. View at: Publisher Site | PubMed
[41] M L Dell'Acqua, M C Faux, J Thorburn,
et al. “Membrane-targeting sequences on AKAP79 bind
phosphatidylinositol-4,5-bisphosphate.” EMBO J, vol. 17, no. 8, pp.
2246-2260, 1998 View at: Publisher
Site | PubMed
[42] T M Klauck, M C Faux, K Labudda, et
al. “Coordination of three signaling enzymes by AKAP79, a mammalian scaffold
protein.” Science, vol. 271, no. 5255, pp. 1589-1592, 1996. View at: Publisher Site | PubMed
[43] Riad Efendiev, Bret K Samelson, Bao T
Nguyen, et al. “AKAP79 interacts with multiple adenylyl cyclase (AC) isoforms
and scaffolds AC5 and -6 to
alpha-amino-3-hydroxyl-5-methyl-4-isoxazole-propionate (AMPA) receptors.” J
Biol Chem, vol. 285, no. 19, pp. 14450-14458, 2010. View at: Publisher Site | PubMed
[44] Debbie Willoughby, Nanako Masada,
Sebastian Wachten, et al. “AKAP79/150 interacts with AC8 and regulates
Ca2+-dependent cAMP synthesis in pancreatic and neuronal systems.” J Biol
Chem, vol. 285, no. 26, pp. 20328-20342, 2010. View at: Publisher Site | PubMed
[45] Dell'Acqua, M. L., K. L. Dodge, S. J.
Tavalin and J. D. Scott (2002). “Mapping the protein phosphatase-2B anchoring
site on AKAP79. Binding and inhibition of phosphatase activity are mediated by
residues 315-360.” J Biol Chem 277(50): 48796-48802. View at: Publisher Site | PubMed
[46] Matthew Watson, Teresa B Almeida,
Arundhati Ray, et al. “Hidden Multivalency in Phosphatase Recruitment by a
Disordered AKAP Scaffold.” J Mol Biol, vol. 434, no. 16, pp. 167682,
2022. View at: Publisher
Site | PubMed
[47] H J Chung, J Xia, R H Scannevin, et
al. “Phosphorylation of the AMPA receptor subunit GluR2 differentially
regulates its interaction with PDZ domain-containing proteins.” J Neurosci,
vol. 20, no. 19, pp. 7258-7267, 2000. View at: Publisher
Site | PubMed
[48] Holly R Robertson, Emily S Gibson,
Timothy A Benke, et al. “Regulation of postsynaptic structure and function by
an A-kinase anchoring protein-membrane-associated guanylate kinase scaffolding
complex.” J Neurosci, vol. 29, no. 24, pp. 7929-7943, 2009. View at: Publisher Site | PubMed
[49] Yuan Lu, Xiang-ming Zha, Eun Young
Kim, et al. “A kinase anchor protein 150 (AKAP150)-associated protein kinase A
limits dendritic spine density.” J Biol Chem, vol. 286, no. 30, pp.
26496-26506, 2011. View at: Publisher
Site | PubMed
[50] Alicia M Purkey, Kevin M Woolfrey,
Kevin C Crosby, et al. “AKAP150 Palmitoylation Regulates Synaptic Incorporation
of Ca(2+)-Permeable AMPA Receptors to Control LTP.” Cell Rep, vol. 25,
no. 4, pp. 974-987.e4, 2018. View at: Publisher Site | PubMed
[51] Jiangchuan Tao, Craig C Malbon
“G-protein-coupled receptor-associated A-kinase anchoring proteins AKAP5 and
AKAP12: differential signaling to MAPK and GPCR recycling.” J Mol Signal,
vol. 3, pp. 19, 2008. View at: Publisher
Site | PubMed
[52] Yanjing Guo, Tao Bo, Xinli Zhou, et
al. “AKAP5 signaling complexes: focal points and functional properties.” Neuro
Endocrinol Lett, vol. 36, no. 1, pp. 7-14, 2015. View at: PubMed
[53] Lisa L Gomez, Shuvo Alam, Karen E
Smith, “Regulation of A-kinase anchoring protein 79 150-cAMP-depen-dent protein
kinase postsynaptic targeting by NMDA receptor activationof calcineurin and
remodeling of dendritic actin.” J Neurosci, vol. 22, no. 16, pp.
7027-7044, 2002. View at: Publisher
Site | PubMed
[54] Jessica A Gorski, Lisa L Gomez, John
D Scott, et al. “Association of an A-kinase-anchoring protein signaling
scaffold with cadherin adhesion molecules in neurons and epithelial cells.” Mol
Biol Cell, vol. 16, no. 8, pp. 3574-3590, 2005. View at: Publisher Site | PubMed
[55] Xenia Gorny, Marina Mikhaylova,
Christian Seeger, et al. “AKAP79/150 interacts with the neuronal
calcium-binding protein caldendrin.” J Neurochem, vol. 122, no. 4, pp.
714-726, 2012. View at: Publisher Site | PubMed
[56] Carmen W Dessauer “Adenylyl
cyclase--A-kinase anchoring protein complexes: the next dimension in cAMP
signaling.” Mol Pharmacol, vol. 76, no. 5, pp. 935-941, 2009. View at: Publisher Site | PubMed
[57] Mingxu Zhang, Tommaso Patriarchi,
Ivar S Stein, et al. “Adenylyl cyclase anchoring by a kinase anchor protein
AKAP5 (AKAP79/150) is important for postsynaptic beta-adrenergic signaling.” J
Biol Chem, vol. 288, no. 24, pp. 17918-17931, 2013. View at: Publisher Site | PubMed
[58] Eric A Horne, Mark L Dell'Acqua
“Phospholipase C is required for changes in postsynaptic structure and function
associated with NMDA receptor-dependent long-term depression.” J Neurosci,
vol. 27, no. 13, pp. 3523-3534, 2007. View at: Publisher Site | PubMed
[59] Xuming Zhang, Lin Li, Peter A McNaughton
“Proinflammatory mediators modulate the heat-activated ion channel TRPV1 via
the scaffolding protein AKAP79/150.” Neuron, vol. 59, no. 3, pp.
450-461, 2008. View at: Publisher
Site | PubMed
[60] Christophe Altier, Stefan J Dubel,
Christian Barrère, et al. “Trafficking of L-type calcium channels mediated by
the postsynaptic scaffolding protein AKAP79.” J Biol Chem, vol. 277, no.
37, pp. 33598-33603, 2002. View at: Publisher Site | PubMed
[61] Jie Zhang, Manjot Bal, Sonya
Bierbower, et al. “AKAP79/150 signal complexes in G-protein modulation of
neuronal ion channels.” J Neurosci, vol. 31, no. 19, pp. 7199-7211,
2011. View at: Publisher
Site | PubMed
[62] Sunghee Chai, Minghua Li, JingQuan
Lan, et al. “A kinase-anchoring protein 150 and calcineurin are involved in
regulation of acid-sensing ion channels ASIC1a and ASIC2a.” J Biol Chem,
vol. 282, no. 31, pp. 22668-22677, 2007. View at: Publisher Site | PubMed
[63] Can Gao, Natalie C Tronson, Jelena
Radulovic “Modulation of behavior by scaffolding proteins of the post-synaptic
density.” Neurobiol Learn Mem, vol. 105, pp. 3-12, 2013. View at: Publisher Site | PubMed
[64] J D Scott “Cyclic
nucleotide-dependent protein kinases.” Pharmacol Ther, vol. 50, no. 1,
pp. 123-145, 1991. View at: Publisher Site | PubMed
[65] F W Herberg, A Maleszka, T Eide,
“Analysis of A-kinase anchoring protein (AKAP) interaction with protein kinase
A (PKA) regulatory subunits: PKA isoform specificity in AKAP binding.” J Mol
Biol, vol. 298, no. 2, pp. 329-339, 2000. View at: Publisher Site | PubMed
[66] J D Scott, M L Dell'Acqua, I D
Fraser, et al. “Coordination of cAMP signaling events through PKA anchoring.” Adv
Pharmacol, vol. 47, pp. 175-207, 2000. View at: Publisher Site | PubMed
[67] M G Newlon, M Roy, D Morikis, et al.
“A novel mechanism of PKA anchoring revealed by solution structures of
anchoring complexes.” EMBO J, vol. 20, no. 7, pp. 1651-1662, 2001. View
at: Publisher Site | PubMed
[68] F Donelson Smith, Jessica L Esseltine
1, Patrick J Nygren. “Local protein kinase A action proceeds through intact
holoenzymes.” Science, vol. 356, no. 6344, pp. 1288-1293, 2017. View at:
Publisher Site | PubMed
[69] Ping Zhang, Eric V Smith-Nguyen,
Malik M Keshwani, et al. “Structure and allostery of the PKA RIIbeta tetrameric
holoenzyme.” Science, vol. 335, no. 6069, pp. 712-716, 2012. View at: Publisher Site | PubMed
[70] Haining Zhong, Gek-Ming Sia, Takashi
R Sato, et al. “Subcellular dynamics of type II PKA in neurons.” Neuron,
vol. 62, no. 3, pp. 363-374, 2009. View at: Publisher Site | PubMed
[71] Alexandra C Newton “Protein kinase C:
perfectly balanced.” Crit Rev Biochem Mol Biol, vol. 53, no. 2, pp. 208-230,
2018. View at: Publisher
Site | PubMed
[72] Takahito Kawano, Junichi Inokuchi 2,
Masatoshi Eto, et al. “Activators and Inhibitors of Protein Kinase C (PKC):
Their Applications in Clinical Trials.” Pharmaceutics, vol. 13, no. 11,
pp. 1748, 2021. View at: Publisher
Site | PubMed
[73] G M Ramakers, P Pasinelli, J J Hens,
“Protein kinase C in synaptic plasticity changes in the in situ phosphorylation
state of identified pre-andpostsynaptic substrates.” Prog
Neuropsychopharmacol Biol Psychiatry, vol. 21, no. 3, pp. 455-486, 1997.
View at: Publisher
Site | PubMed
[74] Erika Abrial, Adeline Etievant,
Cécile Bétry, “Protein kinase C regulates mood-related behaviors and adult
hippocampal cell proliferation in rats.” Prog Neuropsychopharmacol Biol
Psychiatry, vol. 43, pp. 40-48, 2013. View at: Publisher Site | PubMed
[75] M D Ehlers, W G Tingley, R L Huganir
“Regulated subcellular distribution of the NR1 subunit of the NMDA receptor.” Science,
vol. 269, no. 5231, pp. 1734-1737, 1995. View at: Publisher Site | PubMed
[76] Elaine D Por, Bret K Samelson, Sergei
Belugin, et al. “PP2B/calcineurin-mediated desensitization of TRPV1 does not
require AKAP150.” Biochem J, vol. 432, no. 3, pp. 549-556, 2010. View
at: Publisher Site | PubMed
[77] Patrick J Nygren, John D Scott
“Regulation of the phosphatase PP2B by protein-protein interactions.” Biochem
Soc Trans, vol. 44, no. 5, pp. 1313-1319, 2016. View at: Publisher Site | PubMed
[78] Huiming Li, Matthew D Pink, Jonathan
G Murphy, “Balanced interactions of calcineurin with AKAP79 regulate
Ca2+-calcineurin-NFAT signaling.” Nat Struct Mol Biol, vol. 19, no. 3,
pp. 337-345, 2012. View at: Publisher
Site | PubMed
[79] Zhanmin Lin, Bin Wu, Maarten W Paul,
et al. “Protein Phosphatase 2B Dual Function Facilitates Synaptic Integrity and
Motor Learning.” J Neurosci, vol. 41, no. 26, pp. 5579-5594, 2021. View
at: Publisher
Site | PubMed
[80] Timothy W Church, Parul Tewatia, Saad
Hannan, et al. “AKAP79 enables calcineurin to directly suppress protein kinase
A activity.” Elife, vol. 10, pp. e68164, 2021. View at: Publisher Site | PubMed
[81] Matthew G Gold “A frontier in the
understanding of synaptic plasticity: solving the structure of the postsynaptic
density.” Bioessays, vol. 34, no. 7, pp. 599-608, 2012. View at: Publisher Site | PubMed
[82] Joshua L Smalley, Georgina Kontou,
Catherine Choi, et al. “Isolation and Characterization of Multi-Protein
Complexes Enriched in the K-Cl Co-transporter 2 From Brain Plasma Membranes.” Front
Mol Neurosci, vol. 13, pp. 563091, 2020. View at: Publisher Site | PubMed
[83] Karen E Smith, Emily S Gibson, Mark L
Dell'Acqua “cAMP-dependent protein kinase postsynaptic localization regulated
by NMDA receptor activation through translocation of an A-kinase anchoring
protein scaffold protein.” J Neurosci, vol. 26, no. 9, pp. 2391-2402,
2006. View at: Publisher
Site | PubMed
[84] Sandra Jurado, Virginie Biou, Robert
C Malenka “A calcineurin/AKAP complex is required for NMDA receptor-dependent
long-term depression.” Nat Neurosci, vol. 13, no. 9, pp. 1053-1055,
2010. View at: Publisher
Site | PubMed
[85] Jennifer L Sanderson, Jessica A
Gorski, Emily S Gibson, et al. “AKAP150-anchored calcineurin regulates synaptic
plasticity by limiting synaptic incorporation of Ca2+-permeable AMPA
receptors.” J Neurosci, vol. 32, no. 43, pp. 15036-15052, 2012. View at:
Publisher
Site | PubMed
[86] Johannes W Hell “How Ca2+-permeable
AMPA receptors, the kinase PKA, and the phosphatase PP2B are intertwined in
synaptic LTP and LTD.” Sci Signal, vol. 9, no. 425, pp. e2, 2016. View
at: Publisher Site | PubMed
[87] Jennifer L Sanderson, Jessica A
Gorski, Mark L Dell'Acqua “NMDA Receptor-Dependent LTD Requires Transient
Synaptic Incorporation of Ca(2)(+)-Permeable AMPARs Mediated by
AKAP150-Anchored PKA and Calcineurin.” Neuron, vol. 89, no. 5, pp.
1000-1015, 2016. View at: Publisher
Site | PubMed
[88] Jennifer L Sanderson, John D Scott,
Mark L Dell'Acqua “Control of Homeostatic Synaptic Plasticity by AKAP-Anchored
Kinase and Phosphatase Regulation of Ca(2+)-Permeable AMPA Receptors.” J
Neurosci, vol. 38, no. 11, pp. 2863-2876, 2018. View at: Publisher Site | PubMed
[89] Alicia M Purkey, Mark L Dell'Acqua
“Phosphorylation-Dependent Regulation of Ca(2+)-Permeable AMPA Receptors During
Hippocampal Synaptic Plasticity.” Front Synaptic Neurosci, vol. 12, pp.
8, 2020. View at: Publisher
Site | PubMed
[90] B D Johnson, T Scheuer, W A Catterall
“Voltage-dependent potentiation of L-type Ca2+ channels in skeletal muscle
cells requires anchored cAMP-dependent protein kinase.” Proc Natl Acad Sci U
S A, vol. 91, no. 24, pp. 11492-11496, 1994. View at: Publisher Site | PubMed
[91] Richard D Swayze, Marie-France Lisé,
Joshua N Levinson, et al. “Modulation of dopamine mediated phosphorylation of
AMPA receptors by PSD-95 and AKAP79/150.” Neuropharmacology, vol. 47,
no. 5, pp. 764-778, 2004. View at: Publisher Site | PubMed
[92] Y Ben-Ari, L Aniksztejn, P
Bregestovski “Protein kinase C modulation of NMDA currents: an important link
for LTP induction.” Trends Neurosci, vol. 15, no. 9, pp. 333-339, 1992.
View at: Publisher
Site | PubMed
[93] L D Snell, K R Iorio, B Tabakoff, et
al. “Protein kinase C activation attenuates N-methyl-D-aspartate-induced
increases in intracellular calcium in cerebellar granule cells.” J Neurochem,
vol. 62, no. 5, pp. 1783-1789, 1994. View at: Publisher
Site | PubMed
[94] D N Lieberman, I Mody “Regulation of
NMDA channel function byendogenous Ca(2+)-dependent phosphatase.” Nature,
vol. 369, no. 6477, pp. 235-239, 1994. View at: Publisher Site | PubMed
[95] G Tong, D Shepherd, C E Jahr
“Synaptic desensitization of NMDA receptors by calcineurin.” Science,
vol. 267, no. 5203, pp. 1510-1512, 1995. View at: Publisher Site | PubMed
[96] Z E Hausken, J D Scott “Properties of
A-kinase anchoring proteins.” Biochem Soc Trans, vol. 24, no. 4, pp.
986-991, 1996. View at: Publisher
Site | PubMed
[97] T G Banke, D Bowie, H Lee, et al.
“Control of GluR1 AMPA receptor function by cAMP-dependent protein kinase.” J
Neurosci, vol. 20, no. 1, pp. 89-102, 2000. View at: Publisher
Site | PubMed
[98] M D Ehlers “Reinsertion or
Degradation of AMPA Receptors Determined by Activity-Dependent Endocytic
Sorting.” Neuron, vol. 28, no. 2, pp. 511-525, 2000. View at: Publisher Site | PubMed
[99] José A Esteban, Song-Hai Shi,
Christopher Wilson, et al. “PKA phosphorylation of AMPA receptor subunits controls
synaptic trafficking underlying plasticity.” Nat Neurosci, vol. 6, no.
2, pp. 136-143, 2003. View at: Publisher
Site | PubMed
[100] Hey-Kyoung Lee, Kogo Takamiya,
Jung-Soo Han, et al. “Phosphorylation of the AMPA Receptor GluR1 Subunit Is
Required for Synaptic Plasticity and Retention of Spatial Memory.” Cell,
vol. 112, no. 5, pp. 631-643, 2003. View at: Publisher Site | PubMed
[101] Michael C Oh, Victor A Derkach, Eric
S Guire, et al. “Extrasynaptic membrane trafficking regulated by GluR1 serine
845 phosphorylation primes AMPA receptors for long-term potentiation.” J
Biol Chem, vol. 281, no. 2, pp. 752-758, 2006. View at: Publisher Site | PubMed
[102] Heng-Ye Man, Yoko Sekine-Aizawa,
Richard L Huganir “Regulation of
{alpha}-amino-3-hydroxy-5-methyl-4-isoxazolepropionic acid receptor trafficking
through PKA phosphorylation of the Glu receptor 1 subunit.” Proc Natl Acad
Sci U S A, vol. 104, no. 9, pp. 3579-3584, 2007. View at: Publisher Site | PubMed
[103] Graham H Diering, Ahleah S Gustina,
Richard L Huganir “PKA-GluA1 coupling via AKAP5 controls AMPA receptor
phosphorylation and cell-surface targeting during bidirectional homeostatic
plasticity.” Neuron, vol. 84, no. 4, pp. 790-805, 2014. View at: Publisher Site | PubMed
[104] H K Lee, M Barbarosie, K Kameyama, et
al. “Regulation of distinct AMPA receptor phosphorylation sites during
bidirectional synaptic plasticity.” Nature, vol. 405, no. 6789, pp.
955-959, 2000. View at: Publisher
Site | PubMed
[105] Eric M Snyder, Marcie Colledge,
Robert A Crozier, et al. “Role for A Kinase-anchoring Proteins (AKAPS) in
Glutamate Receptor Trafficking and Long Term Synaptic Depression.” J Biol
Chem, vol. 280, no. 17, pp. 16962-16968, 2005. View at: Publisher Site | PubMed
[106] Yuan Lu, Mingxu Zhang, Indra A Lim,
et al. “AKAP150-anchored PKA activity is important for LTD during its induction
phase.” J Physiol, vol. 586, no. 17, pp. 4155-4164, 2008. View at: Publisher Site | PubMed
[107] Wenwen Cheng, Dolores
Siedlecki-Wullich, Judit Català-Solsona, et al. “Proteasomal-Mediated
Degradation of AKAP150 Accompanies AMPAR Endocytosis during cLTD.” eNeuro,
vol. 7, no. 2, pp. ENEURO.0218-19.2020, 2020. View at: Publisher Site
| PubMed
[108] V Derkach, A Barria, T R Soderling
“Ca2+/calmodulin-kinase II enhances channel conductance of
alpha-amino-3-hydroxy-5-methyl-4-isoxazolepropionate type glutamate receptors.”
Proc Natl Acad Sci U S A, vol. 96, no. 6, pp. 3269-3274, 1999. View at: Publisher Site | PubMed
[109] Steven J Tavalin “AKAP79 Selectively
Enhances Protein Kinase C Regulation of GluR1 at a Ca2+-Calmodulin-dependent
Protein Kinase II/Protein Kinase C Site.” J Biol Chem, vol. 283, no. 17,
pp. 11445-11452, 2008. View at: Publisher
Site | PubMed
[110] Anders S Kristensen, Meagan A
Jenkins, Tue G Banke,. “Mechanism of Ca2+/calmodulin-dependent kinase II
regulation of AMPA receptor gating.” Nat Neurosci, vol. 14, no. 6, pp.
727-735, 2011. View at: Publisher
Site | PubMed
[111] Kyle C Summers, Amy S Bogard, Steven
J Tavalin “Preferential generation of Ca(2+)-permeable AMPA receptors by
AKAP79-anchored protein kinase C proceeds via GluA1 subunit phosphorylation at
Ser-831.” J Biol Chem, vol. 294, no. 14, pp. 5521-5535, 2019. View at: Publisher Site | PubMed
[112] Kenneth J Seidenman, Jordan P
Steinberg, Richard Huganir, et al. “Glutamate receptor subunit 2 Serine 880
phosphorylation modulates synaptic transmission and mediates plasticity in CA1
pyramidal cells.” J Neurosci, vol. 23, no. 27, pp. 9220-9228, 2003. View
at: Publisher
Site | PubMed
[113] Jordan P Steinberg, Kogo Takamiya,
Ying Shen,et al. “Targeted in vivo mutations of the AMPA receptor subunit GluR2
and its interacting protein PICK1 eliminate cerebellar long-term depression.” Neuron,
vol. 49, no. 6, pp. 845-860, 2006. View at: Publisher Site | PubMed
[114] Alexis Bavencoffe, Yong Li, Zizhen
Wu, et al. “Persistent Electrical Activity in Primary Nociceptors after Spinal
Cord Injury Is Maintained by Scaffolded Adenylyl Cyclase and Protein Kinase A
and Is Associated with Altered Adenylyl Cyclase Regulation.” J Neurosci,
vol. 36, no. 5, pp. 1660-1668, 2016. View at: Publisher Site | PubMed
[115] Katherine E Brandao, Mark L
Dell'Acqua, S Rock Levinson “A-kinase anchoring protein 150 expression in a
specific subset of TRPV1- and CaV 1.2-positive nociceptive rat dorsal root
ganglion neurons.” J Comp Neurol, vol. 520, no. 1, pp. 81-99, 2012. View
at: Publisher Site | PubMed
[116] D Sarkar, J Erlichman, C S Rubin
“Identification of a calmodulin-binding protein that co-purifies with the
regulatory subunit of brain protein kinase II.” J Biol Chem, vol. 259,
no. 15, pp. 9840-9846, 1984. View at: PubMed
[117] D W Carr, R E Stofko-Hahn, I D
Fraser, et al. “Localization of the cAMP-dependent protein kinase to the
postsynaptic densities by A-kinase anchoring proteins. Characterization of AKAP
79.” J Biol Chem, vol. 267, no. 24, pp. 16816-16823, 1992. View at: PubMed
[118] N Ulfig, M Setzer “Expression of a
kinase anchoring protein 79 in the human fetal amygdala.” Microsc Res Tech,
vol. 46, no. 1, pp. 48-52, 1999. View at: Publisher
Site | PubMed
[119] A Sík, A Gulácsi, Y Lai, et al.
“Localization of the A kinase anchoring protein AKAP79 in the human
hippocampus.” Eur J Neurosci, vol. 12, no. 4, pp. 1155-1164, 2000. View
at: Publisher
Site | PubMed
[120] N Ulfig, F Neudörfer, J Bohl
“Development-related expression of AKAP79 in the striatal compartments of the
human brain.” Cells Tissues Organs, vol. 168, no. 4, pp. 319-329, 2001.
View at: Publisher Site | PubMed
[121] Marta A P Moita, Raphael Lamprecht,
Karim Nader, et al. “A-kinase anchoring proteins in amygdala are involved in
auditory fear memory.” Nat Neurosci, vol. 5, no. 9, pp. 837-838, 2002.
View at: Publisher Site | PubMed
[122] Simon A Hinke, Manuel F Navedo,
Allison Ulman, et al. “Anchored phosphatases modulate glucose homeostasis.” EMBO
J, vol. 31, no. 20, pp. 3991-4004, 2012. View at: Publisher Site | PubMed
[123] C Blake Nichols, Charles F Rossow,
Manuel F Navedo, et al. “Sympathetic stimulation of adult cardiomyocytes
requires association of AKAP5 with a subpopulation of L-type calcium channels.”
Circ Res, vol. 107, no. 6, pp. 747-756, 2010. View at: Publisher Site | PubMed
[124] Manuel F Navedo, Madeline
Nieves-Cintrón, Gregory C Amberg, et al. “AKAP150 is required for stuttering
persistent Ca2+ sparklets and angiotensin II-induced hypertension.” Circ Res,
vol. 102, no. 2, pp. e1-e11, 2008. View at: Publisher Site | PubMed
[125] Madeline Nieves-Cintrón, Gregory C
Amberg, Manuel F Navedo, et al. “The control of Ca2+ influx and NFATc3
signaling in arterial smooth muscle during hypertension.” Proc Natl Acad Sci
U S A, vol. 105, no. 40, pp. 15623-15628, 2008. View at: Publisher Site | PubMed
[126] K L Dodge, D W Carr, C Yue, et al. “A
role for AKAP (A kinase anchoring protein) scaffolding in the loss of a cyclic
adenosine 3',5'-monophosphate inhibitory response in late pregnant rat
myometrium.” Mol Endocrinol, vol. 13, no. 12, pp. 1977-1987, 1999. View
at: Publisher Site | PubMed
[127] G Xie, J P Raufman “Association of
protein kinase A with AKAP150 facilitates pepsinogen secretion from gastric
chief cells.” Am J Physiol Gastrointest Liver Physiol, vol. 281, no. 4,
pp. G1051-G1058, 2001. View at: Publisher Site | PubMed
[128] K Kurihara, N Nakanishi “Regulation
of Na,K-ATPase by cAMP-dependent protein kinase anchored on membrane via
A-kinase anchoring protein subtype, AKAP-150, in rat parotid gland.” Ann N Y
Acad Sci, vol. 986, pp. 636-638, 2003. View at: Publisher Site | PubMed
[129] Ching-Yi Wu, Dennis H DiJulio, Kerry
L Jacobson, et al. “The contribution of AKAP5 in amylase secretion from mouse
parotid acini.” Am J Physiol Cell Physiol, vol. 298, no. 5, pp.
C1151-C1158, 2010. View at: Publisher
Site | PubMed
[130] Ji-Heon Rhim, Ik-Soon Jang, Eui-Ju
Yeo, et al. “Role of protein kinase C-dependent A-kinase anchoring proteins in
lysophosphatidic acid-induced cAMP signaling in human diploid fibroblasts.” Aging
Cell, vol. 5, no. 6, pp. 451-461, 2006. View at: Publisher Site | PubMed
[131] Jiangwen Lu, Wangsheng Wang 1 2,
Yabing Mi, et al. “AKAP95-mediated nuclear anchoring of PKA mediates
cortisol-induced PTGS2 expression in human amnion fibroblasts.” Sci Signal,
vol. 10, no. 506, pp. eaac6160, 2017. View at: Publisher Site | PubMed
[132] Robynn V Schillace, Sarah F Andrews,
Greg A Liberty, et al. “Identification and characterization of myeloid
translocation gene 16b as a novel a kinase anchoring protein in T lymphocytes.”
J Immunol, vol. 168, no. 4, pp. 1590-1599, 2002. View at: Publisher Site | PubMed
[133] N Ulfig, W Y Chan “Expression of a
kinase anchoring protein 79 and synaptophysin in the developing human red
nucleus.” Neurosignals, vol. 11, no.2, pp. 95-102, 2002. View at: Publisher Site | PubMed
[134] A Feliciello, M E Gottesman, E V
Avvedimento “The biological functions of A-kinase anchor proteins.” J Mol
Biol, vol. 308, no. 2, pp. 99-114, 2001. View at: Publisher Site | PubMed
[135] Anghelus Ostroveanu, Eddy A Van der
Zee, Amalia M Dolga, et al. “A-kinase anchoring protein 150 in the mouse brain
is concentrated in areas involved in learning and memory.” Brain Res,
vol. 1145, pp. 97-107, 2007. View at: Publisher Site | PubMed
[136] Maurine E Linder, Robert J Deschenes
“Palmitoylation: policing protein stability and traffic.” Nat Rev Mol Cell
Biol, vol. 8, no. 1, pp. 74-84, 2007. View at: Publisher Site | PubMed
[137] Yuko Fukata, Masaki Fukata “Protein
palmitoylation in neuronal development and synaptic plasticity.” Nat Rev
Neurosci, vol. 11, no. 3, pp. 161-175, 2010. View at: Publisher Site | PubMed
[138] Robbins Puthenveetil, Natalia
Gómez-Navarro, Anirban Banerjee “Access and utilization of long chain fatty
acyl-CoA by zDHHC protein acyltransferases.” Curr Opin Struct Biol, vol.
77, pp. 102463, 2022. View at: Publisher Site | PubMed
[139] Kun Huang, Anat Yanai, Rujun Kang, et
al. “Huntingtin-interacting protein HIP14 is a palmitoyl transferase involved
in palmitoylation and trafficking of multiple neuronal proteins.” Neuron,
vol. 44, no. 6, pp. 977-986, 2004. View at: Publisher Site | PubMed
[140] John T Swarthout, Sandra Lobo, Lynn
Farh, et al. “DHHC9 and GCP16 constitute a human protein fatty acyltransferase
with specificity for H- and N-Ras.” J Biol Chem, vol. 280, no. 35, pp.
31141-31148, 2005. View at: Publisher
Site | PubMed
[141] Heesung Sohn, Mikyoung Park
“Palmitoylation-mediated synaptic regulation of AMPA receptor trafficking and
function.” Arch Pharm Res, vol. 42, no. 5, pp. 426-435, 2019. View at: Publisher Site | PubMed
[142] Tamal Sadhukhan, Maria B Bagh,
Abhilash P Appu, et al. “In a mouse model of INCL reduced S-palmitoylation of
cytosolic thioesterase APT1 contributes to microglia proliferation and
neuroinflammation.” J Inherit Metab Dis, vol. 44, no. 4, pp. 1051-1069,
2021. View at: Publisher
Site | PubMed
[143] Jennifer Greaves, Juliet A
Carmichael, Luke H Chamberlain “The palmitoyl transferase DHHC2 targets a
dynamic membrane cycling pathway: regulation by a C-terminal domain.” Mol
Biol Cell, vol. 22, no. 11, pp. 1887-1895, 2011. View at: Publisher Site | PubMed
[144] Christine Salaun, Louise Ritchie 1,
Jennifer Greaves, et al. “The C-terminal domain of zDHHC2 contains distinct
sorting signals that regulate intracellular localisation in neurons and
neuroendocrine cells.” Mol Cell Neurosci, vol. 85, pp. 235-246, 2017.
View at: Publisher
Site | PubMed
[145] Han Zhang, Xiuli Li, Chuanchuan Ma,
et al. “Fine-mapping of ZDHHC2 identifies risk variants for schizophrenia in
the Han Chinese population.” Mol Genet Genomic Med, vol. 8, no. 7, pp.
e1190, 2020. View at: Publisher
Site | PubMed
[146] Kevin M Woolfrey, Jennifer L
Sanderson, Mark L Dell'Acqua “The palmitoyl acyltransferase DHHC2 regulates
recycling endosome exocytosis and synaptic potentiation through palmitoylation
of AKAP79/150.” J Neurosci, vol. 35, no. 2, pp. 442-456, 2015. View at: Publisher Site | PubMed
[147] Ilse Delint-Ramirez, Debbie
Willoughby, Gerald R V Hammond,. “Palmitoylation targets AKAP79 protein to
lipid rafts and promotes its regulation of calcium-sensitive adenylyl cyclase
type 8.” J Biol Chem, vol. 286, no. 38, pp. 32962-32975, 2011. View at: Publisher Site | PubMed
[148] Jun Han, Pengfei Wu, Fang Wang, et al
“S-palmitoylation regulates AMPA receptors trafficking and function: a novel
insight into synaptic regulation and therapeutics.” Acta Pharm Sin B,
vol. 5, no. 1, pp. 1-7, 2015. View at: Publisher Site | PubMed
[149] Xiaobing Chen, Kevin C Crosby, Austin
Feng, et al. “Palmitoylation of A-kinase anchoring protein 79/150 modulates its
nanoscale organization, trafficking, and mobility in postsynaptic spines.” Front
Synaptic Neurosci, vol. 14, pp. 1004154, 2022. View at: Publisher Site | PubMed
[150] Jennifer L Sanderson, Ronald K
Freund, Jessica A Gorski “β-Amyloid disruption of LTP/LTD balance is mediated by
AKAP150-anchored PKA and Calcineurin regulation of Ca(2+)-permeable AMPA
receptors.” Cell Rep,
vol. 37, no. 1, pp. 109786, 2021. View at: Publisher Site | PubMed
[151] Anton Omelchenko, Bonnie L Firestein
“Lipids and phosphates at odds in synaptic depression.” J Biol Chem,
vol. 293, no. 5, pp. 1568-1569, 2018. View at: Publisher Site | PubMed
[152] Kevin M Woolfrey, Heather O'Leary,
Dayton J Goodell, et al. “CaMKII regulates the depalmitoylation and synaptic
removal of the scaffold protein AKAP79/150 to mediate structural long-term
depression.” J Biol Chem, vol. 293, no. 5, pp. 1551-1567, 2018. View at:
Publisher Site | PubMed
[153] Katherine A Mifflin, Bradley J Kerr
“The transition from acute to chronic pain: understanding how different
biological systems interact.” Can J Anaesth, vol. 61, no. 2, pp.
112-122, 2014. View at: Publisher
Site | PubMed
[154] G Bhave, F Karim, S M Carlton, et al.
“Peripheral group I metabotropic glutamate receptors modulate nociception in
mice.” Nat Neurosci, vol. 4, no. 4, pp. 417-423, 2001. View at: Publisher Site | PubMed
[155] Gábor Petho, Peter W Reeh “Sensory
and signaling mechanisms of bradykinin, eicosanoids, platelet-activating
factor, and nitric oxide in peripheral nociceptors.” Physiol Rev, vol.
92, no. 4, pp. 1699-1775, 2012. View at: Publisher Site | PubMed
[156] Ru-Rong Ji, Zhen-Zhong Xu, Yong-Jing
Gao “Emerging targets in neuroinflammation-driven chronic pain.” Nat Rev
Drug Discov, vol. 13, no. 7, pp. 533-548, 2014. View at: Publisher Site | PubMed
[157] Pankaj Baral, Swalpa Udit, Isaac M
Chiu “Pain and immunity: implications for host defence.” Nat Rev Immunol,
vol. 19, no. 7, pp. 433-447, 2019. View at: Publisher Site | PubMed
[158] Salman Khan, Omer Shehzad, Jaemoo
Chun, et al. “Mechanism underlying anti-hyperalgesic and anti-allodynic
properties of anomalin in both acute and chronic inflammatory pain models in
mice through inhibition of NF-kappaB, MAPKs and CREB signaling cascades.” Eur
J Pharmacol, vol. 718, no. 1-3, pp. 448-458, 2013. View at: Publisher Site | PubMed
[159] Stuart Bevan, Talisia Quallo, David A
Andersson “Trpv1.” Handb Exp Pharmacol, vol. 222, pp. 207-245, 2014.
View at: Publisher
Site | PubMed
[160] Parvinder Kaur Rathee, Carsten
Distler, Otilia Obreja, et al. “PKA/AKAP/VR-1 module: A common link of
Gs-mediated signaling to thermal hyperalgesia.” J Neurosci, vol. 22, no.
11, pp. 4740-4745, 2002. View at: Publisher
Site | PubMed
[161] Nathaniel A Jeske, Anibal Diogenes,
Nikita B Ruparel, et al. “A-kinase anchoring protein mediates TRPV1 thermal
hyperalgesia through PKA phosphorylation of TRPV1.” Pain, vol. 138, no.
3, pp. 604-616, 2008. View at: Publisher Site | PubMed
[162] Nathaniel A Jeske, Amol M Patwardhan,
Nikita B Ruparel, et al. “A-kinase anchoring protein 150 controls protein
kinase C-mediated phosphorylation and sensitization of TRPV1.” Pain,
vol. 146, no. 3, pp. 301-307, 2009. View at: Publisher Site | PubMed
[163] Katrin Schnizler, Leonid P Shutov,
Michael J Van Kanegan, et al. “Protein kinase A anchoring via AKAP150 is
essential for TRPV1 modulation by forskolin and prostaglandin E2 in mouse
sensory neurons.” J Neurosci, vol. 28, no. 19, pp. 4904-4917, 2008. View
at: Publisher
Site | PubMed
[164] Jongseok Lee, Man-Kyo Chung, Jin Y Ro
“Activation of NMDA receptors leads to phosphorylation of TRPV1 S800 by protein
kinase C and A-Kinase anchoring protein 150 in rat trigeminal ganglia.” Biochem
Biophys Res Commun, vol. 424, no. 2, pp. 358-363, 2012. View at: Publisher Site | PubMed
[165] Michael J M Fischer, Joan Btesh,
Peter A McNaughton “Disrupting Sensitization of Transient Receptor Potential
Vanilloid Subtype 1 Inhibits Inflammatory Hyperalgesia.” J Neurosci,
vol. 33, no. 17, pp. 7407-7414, 2013. View at: Publisher Site | PubMed
[166] Man-Kyo Chung, Jongseok Lee, John
Joseph, et al. “Peripheral group I metabotropic glutamate receptor activation
leads to muscle mechanical hyperalgesia through TRPV1 phosphorylation in the
rat.” J Pain, vol. 16, no. 1, pp. 67-76, 2015. View at: Publisher Site | PubMed
[167] Joan Btesh, Michael J M Fischer,
Katherine Stott, et al. “Mapping the Binding Site of TRPV1 on AKAP79:
Implications for Inflammatory Hyperalgesia.” J Neurosci, vol. 33, no.
21, pp. 9184-9193, 2013. View at: Publisher Site | PubMed
[168] Nathaniel A Jeske, Elaine D Por,
Sergei Belugin, Sraboni Chaudhury, et al. “A-kinase anchoring protein 150
mediates transient receptor potential family V type 1 sensitivity to
phosphatidylinositol-4,5-bisphosphate.” J Neurosci, vol. 31, no. 23, pp.
8681-8688, 2011. View at: Publisher Site | PubMed
[169] Kalina Szteyn, Matthew P Rowan, Ruben
Gomez, et al “A-kinase anchoring protein 79/150 coordinates metabotropic
glutamate receptor sensitization of peripheral sensory neurons.” Pain,
vol. 156, no. 11, pp. 2364-2372, 2015. View at: Publisher Site | PubMed
[170] Ruben Gomez, Dorothy M Kohler,
Allison D Brackley, et al. “Serum response factor mediates nociceptor
inflammatory pain plasticity.” Pain Rep, vol. 3, no. 3, pp. e658, 2018.
View at: Publisher
Site | PubMed
[171] Yinxia Li, Xue Bai, Min Gao, et al.
“AKAP150 and its Palmitoylation Contributed to Pain Hypersensitivity Via
Facilitating Synaptic Incorporation of GluA1-Containing AMPA Receptor in Spinal
Dorsal Horn.” Mol Neurobiol, vol. 58, no. 12, pp. 6505-6519, 2021. View
at: Publisher
Site | PubMed
[172] C Distler, P K Rathee, K S Lips,
“Fast Ca2+-induced potentiation of heat-activated ionic currents requires
cAMP/PKA signaling and functional AKAP anchoring.” J Neurophysiol, vol.
89, no. 5, pp. 2499-2505, 2003. View at: Publisher Site | PubMed
[173] Allan I Basbaum, Diana M Bautista,
Grégory Scherrer, et al. “Cellular and molecular mechanisms of pain.” Cell,
vol. 139, no. 2, pp. 267-284, 2009. View at: Publisher Site | PubMed
[174] Rohini Kuner, Herta Flor “Structural
plasticity and reorganisation in chronic pain.” Nat Rev Neurosci, vol.
18, no. 1, pp. 20-30, 2016. View at: Publisher Site | PubMed
[175] Riad Efendiev, Alexis Bavencoffe,
Hongzhen Hu, et al. “Scaffolding by A-kinase anchoring protein enhances
functional coupling between adenylyl cyclase and TRPV1 channel.” J Biol Chem,
vol. 288, no. 6, pp. 3929-3937, 2013. View at: Publisher Site | PubMed
[176] Shuang Qiu, Ming Zhang, Yan Liu, et
al. “GluA1 phosphorylation contributes to postsynaptic amplification of
neuropathic pain in the insular cortex.” J Neurosci, vol. 34, no. 40,
pp. 13505-13515, 2014. View at: Publisher Site | PubMed
[177] Seth F Oliveria, Mark L Dell'Acqua,
William A Sather “AKAP79/150 anchoring of calcineurin controls neuronal L-type
Ca2+ channel activity and nuclear signaling.” Neuron, vol. 55, no. 2,
pp. 261-275, 2007. View at: Publisher Site | PubMed
[178] Jörg Isensee, Marianne van Cann,
Patrick Despang, et al. “Depolarization induces nociceptor sensitization by
CaV1.2-mediated PKA-II activation.” J Cell Biol, vol. 220, no. 10, pp.
e202002083, 2021.. View at: Publisher
Site | PubMed
[179] Sraboni Chaudhury, Manjot Bal, Sergei
Belugin, et al. “AKAP150-mediated TRPV1 sensitization is disrupted by
calcium/calmodulin.” Mol Pain, vol. 7, pp. 34, 2011. View at: Publisher Site | PubMed
[180] Mitsuko Numazaki, Tomoko Tominaga,
Kumiko Takeuchi, et al. “Structural determinant of TRPV1 desensitization
interacts with calmodulin.” Proc Natl Acad Sci U S A, vol. 100, no. 13,
pp. 8002-8006, 2003. View at: Publisher
Site | PubMed
[181] Fumiko Sekiguchi, Yuka Aoki, Maiko
Nakagawa, et al. “AKAP-dependent sensitization of Ca(v) 3.2 channels via the
EP(4) receptor/cAMP pathway mediates PGE(2) -induced mechanical hyperalgesia.” Br
J Pharmacol, vol. 168, no. 3, pp. 734-745,2013. View at: Publisher Site | PubMed
[182] Hueng-Chuen Fan, Xuming Zhang, Peter
A McNaughton “Activation of the TRPV4 ion channel is enhanced by
phosphorylation.” J Biol Chem, vol. 284, no. 41, pp. 27884-27891, 2009.
View at: Publisher
Site | PubMed
[183] Konrad Mack, Michael J M Fischer
“Disrupting sensitization of TRPV4.” Neuroscience, vol. 352, pp. 1-8,
2017. View at: Publisher
Site | PubMed
[184] Bilin Nie, Cuicui Liu, Xiaohui Bai,
et al. “AKAP150 involved in paclitaxel-induced neuropathic pain via inhibiting
CN/NFAT2 pathway and downregulating IL-4.” Brain Behav Immun, vol. 68,
pp. 158-168, 2018. View at: Publisher
Site | PubMed
[185] Kanako Miyano, Seiji Shiraishi,
Koichiro Minami, et al. “Carboplatin Enhances the Activity of Human Transient
Receptor Potential Ankyrin 1 through the Cyclic AMP-Protein Kinase A-A-Kinase
Anchoring Protein (AKAP) Pathways.” Int J Mol Sci, vol. 20, no. 13, pp.
3271, 2019. View at: Publisher
Site | PubMed
[186] Pablo M Casillas-Espinosa, Kim L
Powell, Terence J O'Brien “Regulators of synaptic transmission: roles in the
pathogenesis and treatment of epilepsy.” Epilepsia, vol. 53 Suppl 9, pp.
41-58, 2012. View at: Publisher
Site | PubMed
[187] Alan Talevi “Antiseizure medication
discovery: Recent and future paradigm shifts.” Epilepsia Open, vol. 7
Suppl 1, no. Suppl 1, pp. S133-S141, 2022. View at: Publisher Site | PubMed
[188] Jessica J Falco-Walter, Ingrid E
Scheffer, Robert S Fisher “The new definition and classification of seizures
and epilepsy.” Epilepsy Res, vol. 139, pp. 73-79, 2018. View at: Publisher Site | PubMed
[189] Gábor Zsurka, Wolfram S Kunz
“Mitochondrial dysfunction and seizures: the neuronal energy crisis.” Lancet Neurol,
vol. 14, no. 9, pp. 956-966, 2015. View at: Publisher Site | PubMed
[190] Mark Manford “Recent advances in
epilepsy.” J Neurol, vol. 264, no. 8, pp. 1811-1824, 2017. View at: Publisher Site | PubMed
[191] Albert Lim, Rhys H Thomas “The
mitochondrial epilepsies.” Eur J Paediatr Neurol, vol. 24, pp. 47-52,
2020. View at: Publisher
Site | PubMed
[192] Hao Deng, Xiaofei Xiu, Zhi Song “The
molecular biology of genetic-based epilepsies.” Mol Neurobiol, vol. 49,
no. 1, pp. 352-367, 2014. View at: Publisher Site | PubMed
[193] Brian J Tunquist, Naoto Hoshi, Eric S
Guire, et al. “Loss of AKAP150 perturbs distinct neuronal processes in mice.” Proc
Natl Acad Sci U S A, vol. 105, no. 34, pp. 12557-12562, 2008. View at: Publisher Site | PubMed
[194] Jie Zhang, Mark S Shapiro.
“Mechanisms and dynamics of AKAP79/150-orchestrated multi-protein signalling
complexes in brain and peripheral nerve.” J Physiol, vol. 594, no. 1, pp.
31-37, 2016. View at: Publisher
Site | PubMed
[195] M C Faux, J D Scott “Regulation of
the AKAP79-protein kinase C interaction by Ca2+/Calmodulin.” J Biol Chem,
vol. 272, no. 27, pp. 17038-17044, 1997. View at: Publisher Site | PubMed
[196] Naoto Hoshi, Jia-Sheng Zhang, Miho
Omaki, et al. “AKAP150 signaling complex promotes suppression of the M-current
by muscarinic agonists.” Nat Neurosci, vol. 6, no. 6, pp. 564-571, 2003.
View at: Publisher Site | PubMed
[197] Haruhiro Higashida, Naoto Hoshi,
Jia-Sheng Zhang, et al. “Protein kinase C bound with A-kinase anchoring protein
is involved in muscarinic receptor-activated modulation of M-type KCNQ
potassium channels.” Neurosci Res, vol. 51, no. 3, pp. 231-234, 2005.
View at: Publisher
Site | PubMed
[198] Eamonn J Dickson, Björn H
Falkenburger, Bertil Hille “Quantitative properties and receptor reserve of the
IP(3) and calcium branch of G(q)-coupled receptor signaling.” J Gen Physiol,
vol. 141, no. 5, pp. 521-535, 2013. View at: Publisher Site | PubMed
[199] Naoto Hoshi “M-Current Suppression,
Seizures and Lipid Metabolism: A Potential Link Between Neuronal Kv7 Channel
Regulation and Dietary Therapies for Epilepsy.” Front Physiol, vol. 11,
pp. 513, 2020. View at: Publisher
Site | PubMed
[200] A Kashishian, M Howard, C Loh, et al.
“AKAP79 inhibits calcineurin through a site distinct from the
immunophilin-binding region.” J Biol Chem, vol. 273, no. 42, pp.
27412-27419, 1998. View at: Publisher
Site | PubMed
[201] Shangli Cai, Shucai Huang, Wei Hao
“New hypothesis and treatment targets of depression: an integrated view of key
findings.” Neurosci Bull, vol. 31, no. 1, pp. 61-74, 2015. View at: Publisher Site | PubMed
[202] Ja Wook Koo, Dipesh Chaudhury,
Ming-Hu Han “Role of Mesolimbic Brain-Derived Neurotrophic Factor in
Depression.” Biol Psychiatry, vol. 86, no. 10, pp. 738-748, 2019. View
at: Publisher
Site | PubMed
[203] J J Schildkraut, P R Draskoczy, E S
Gershon, “Catecholamine metabolism in affective disorders. IV. Preliminary
studies of norepinephrine metabolism in depressed patients treated with
amitriptyline.” J Psychiatr Res, vol. 9, no. 3, pp. 173-185, 1972. View
at: Publisher
Site | PubMed
[204] M Maes, E Vandoolaeghe, R Ranjan,
“Increased serum interleukin-1-receptor-antagonist concentrations in major
depression.” J Affect Disord, vol. 36, no. (1-2), pp. 29-36, 1995. View
at: Publisher
Site | PubMed
[205] R S Duman, J Malberg, J Thome “Neural
plasticity to stress and antidepressant treatment.” Biol Psychiatry,
vol. 46, no. 9, pp. 1181-1191, 1999. View at: Publisher Site | PubMed
[206] Patricia A Zunszain, Christoph
Anacker, Annamaria Cattaneo, et al. “Glucocorticoids, cytokines and brain
abnormalities in depression.” Prog Neuropsychopharmacol Biol Psychiatry,
vol. 35, no. 3, pp. 722-729, 2011. View at: Publisher Site | PubMed
[207] John R Kelly, Yuliya Borre, Ciaran O'
Brien, et al. “Transferring the blues: Depression-associated gut microbiota
induces neurobehavioural changes in the rat.” J Psychiatr Res, vol. 82,
pp. 109-118, 2016. View at: Publisher Site | PubMed
[208] Hanjie Yu, Dan Lv, Mengxin Shen, et
al. “BDNF mediates the protective effects of scopolamine in reserpine-induced
depression-like behaviors via up-regulation of 5-HTT and TPH1.” Psychiatry
Res, vol. 271, pp. 328-334, 2019. View at: Publisher Site | PubMed
[209] Christopher Pittenger, Ronald S Duman
“Stress, depression, and neuroplasticity: a convergence of mechanisms.” Neuropsychopharmacology,
vol. 33, no. 1, pp. 88-109, 2008. View at: Publisher Site | PubMed
[210] Sergio D Iñiguez, Antonio Aubry 2,
Lace M Riggs, et al. “Social defeat stress induces depression-like behavior and
alters spine morphology in the hippocampus of adolescent male C57BL/6 mice.” Neurobiol
Stress, vol. 5, pp. 54-64, 2016. View at: Publisher Site | PubMed
[211] Yu Zhang, Feng Shao 2, Qiong Wang, et
al. “Neuroplastic Correlates in the mPFC Underlying the Impairment of
Stress-Coping Ability and Cognitive Flexibility in Adult Rats Exposed to
Chronic Mild Stress during Adolescence.” Neural Plast, vol. 2017, pp.
9382797, 2017. View at: Publisher
Site | PubMed
[212] Hiroyuki Koike, Michihiko Iijima,
Shigeyuki Chaki “Involvement of AMPA receptor in both the rapid and sustained
antidepressant-like effects of ketamine in animal models of depression.” Behav
Brain Res, vol. 224, no. 1, pp. 107-111, 2011. View at: Publisher Site | PubMed
[213] Byung Kook Lim, Kee Wui Huang, Brad A
Grueter, et al. “Anhedonia requires MC4R-mediated synaptic adaptations in
nucleus accumbens.” Nature, vol. 487, no. 7406, pp. 183-189, 2012. View
at: Publisher Site | PubMed
[214] Eunice Y Yuen, Jing Wei, Wenhua Liu,
“Repeated stress causes cognitive impairment by suppressing glutamate receptor
expression and function in prefrontal cortex.” Neuron, vol. 73, no. 5,
pp. 962-977, 2012. View at: Publisher Site | PubMed
[215] Xiang Cai, Angy J Kallarackal, Mark D
Kvarta, et al. “Local potentiation of excitatory synapses by serotonin and its
alteration in rodent models of depression.” Nat Neurosci, vol. 16, no.
4, pp. 464-472, 2013. View at: Publisher
Site | PubMed
[216] M Gómez-Galán, D De Bundel, A Van
Eeckhaut, et al. “Dysfunctional astrocytic regulation of glutamate transmission
in a rat model of depression.” Mol Psychiatry, vol. 18, no. 5, pp.
582-594, 2013. View at: Publisher
Site | PubMed
[217] Gary M Wilson, Stephane Flibotte,
Vikramjit Chopra, “DNA copy-number analysis in bipolar disorder and schizophrenia
reveals aberrations in genes involved in glutamate signaling.” Hum Mol Genet,
vol. 15, no. 5, pp. 743-749, 2006. View at: Publisher Site | PubMed
[218] Smitha R Sutrala, Dirk Goossens,
Nigel M Williams, et al. “Gene copy number variation in schizophrenia.” Schizophr
Res, vol. 96, no. 1-3, pp. 93-99, 2007. View at: Publisher Site | PubMed
[219] Hans-Gert Bernstein, Henrik
Dobrowolny, Björn H Schott, et al. “Increased density of AKAP5-expressing
neurons in the anterior cingulate cortex of subjects with bipolar disorder.” J
Psychiatr Res, vol. 47, no. 6, pp. 699-705, 2013. View at: Publisher Site | PubMed
[220] G Poelmans, B Franke, D L Pauls, et
al. “AKAPs integrate genetic findings for autism spectrum disorders.” Transl
Psychiatry, vol. 3, no. 6, pp. e270, 2013. View at: Publisher Site | PubMed
[221] Alexandre Surget, Yingjie Wang,
Samuel Leman, et al. “Corticolimbic transcriptome changes are state-dependent
and region-specific in a rodent model of depression and of antidepressant
reversal.” Neuropsychopharmacology, vol. 34, no. 6, pp. 1363-1380, 2009.
View at: Publisher Site | PubMed
[222] Hai-Yun Zhou, Jin-Gang He 2,
Zhuang-Li Hu, et al. “A-Kinase Anchoring Protein 150 and Protein Kinase A
Complex in the Basolateral Amygdala Contributes to Depressive-like Behaviors
Induced by Chronic Restraint Stress.” Biol Psychiatry, vol. 86, no. 2,
pp. 131-142, 2019. View at: Publisher Site | PubMed
[223] Kui Chen, Yu An, Lu Tie, et al.
“Curcumin Protects Neurons from Glutamate-Induced Excitotoxicity by Membrane
Anchored AKAP79-PKA Interaction Network.” Evid Based Complement Alternat Med,
vol. 2015, pp. 706207, 2015. View at: Publisher Site | PubMed
[224] Jean-Claude Béïque, Da-Ting Lin,
Myoung-Goo Kang, et al. “Synapse-specific regulation of AMPA receptor function
by PSD-95.” Proc Natl Acad Sci U S A, vol. 103, no. 51, pp. 19535-19540,
2006. View at: Publisher
Site | PubMed
[225] L B Lester, M C Faux, J B Nauert, et
al. “Targeted protein kinase A and PP-2B regulate insulin secretion through
reversible phosphorylation.” Endocrinology, vol. 142, no. 3, pp.
1218-1227, 2001. View at: Publisher
Site | PubMed
[226] D D Gutterman, M J Durand “Vascular
dysfunction in diabetes mellitus: large conductance calcium-activated potassium
channels as part of a subsarcolemmal signaling soiree.” Circ Res, vol.
114, no. 4, pp. 588-590, 2014. View at: Publisher Site | PubMed
[227] Matthew A Nystoriak, Madeline
Nieves-Cintrón, Patrick J Nygren, et al. “AKAP150 contributes to enhanced
vascular tone by facilitating large-conductance Ca2+-activated K+ channel
remodeling in hyperglycemia and diabetes mellitus.” Circ Res, vol. 114,
no. 4, pp. 607-615, 2014. View at: Publisher Site | PubMed
[228] Madeline Nieves-Cintrón, Matthew A
Nystoriak, Maria Paz Prada, et al. “Selective down-regulation of KV2.1 function
contributes to enhanced arterial tone during diabetes.” J Biol Chem,
vol. 290, no. 12, pp. 7918-7929, 2015. View at: Publisher Site | PubMed
[229] Duane D Hall, Monika A Davare, Mei
Shi, et al. “Critical role of cAMP-dependent protein kinase anchoring to the
L-type calcium channel Cav1.2 via A-kinase anchor protein 150 in neurons.” Biochemistry,
vol. 46, no. 6, pp. 1635-1646, 2007. View at: Publisher Site | PubMed
[230] Matthew A Nystoriak, Madeline
Nieves-Cintrón, Tommaso Patriarchi, et al. “Ser1928 phosphorylation by PKA
stimulates the L-type Ca2+ channel CaV1.2 and vasoconstriction during acute
hyperglycemia and diabetes.” Sci Signal, vol. 10, no. 463, pp. eaaf9647,
2017. View at: Publisher
Site | PubMed
[231] Maria Paz Prada, Arsalan U Syed,
Gopireddy R Reddy, et al. “AKAP5 complex facilitates purinergic modulation of
vascular L-type Ca(2+) channel Ca(V)1.2.” Nat Commun, vol. 11, no. 1,
pp. 5303, 2020. View at: Publisher
Site | PubMed
[232] Miguel Martín-Aragón Baudel, Junyoung
Hong, Johannes W Hell, et al. “Mechanisms of Vascular Ca(V)1.2 Channel
Regulation During Diabetic Hyperglycemia.” Handb Exp Pharmacol. View at:
Publisher Site | PubMed
[233] Lei Li, Jing Li, Benjamin M Drum, et
al. “Loss of AKAP150 promotes pathological remodelling and heart failure
propensity by disrupting calcium cycling and contractile reserve.” Cardiovasc
Res, vol. 113, no. 2, pp. 147-159, 2017. View at: Publisher Site | PubMed
[234] Feng Zhu, Qiushu Wang, Zhi Wang, et
al. “Metoprolol Mitigates Ischemic Heart Remodeling and Fibrosis by Increasing
the Expression of AKAP5 in Ischemic Heart.” Oxid Med Cell Longev, vol.
2022, pp. 5993459, 2022. View at: Publisher
Site | PubMed
[235] T Taigen, L J De Windt, H W Lim, et
al. “Targeted inhibition of calcineurin prevents agonist-induced cardiomyocyte
hypertrophy.” Proc Natl Acad Sci U S A, vol. 97, no. 3, pp. 1196-1201,
2000. View at: Publisher
Site | PubMed
[236] L A Leinwand “Calcineurin inhibition
and cardiac hypertrophy a matter of balance.” Proc Natl Acad Sci U S A,
vol. 98, no. 6, pp. 2947-2949, 2001. View at: Publisher Site | PubMed
[237] A Yatani, R Honda, K M Tymitz, et al.
“Enhanced Ca2+ channel currents in cardiac hypertrophy induced by activation of
calcineurin-dependent pathway.” J Mol Cell Cardiol, vol. 33, no. 2, pp.
249-259, 2001. View at: Publisher
Site | PubMed
[238] Weiguo Zhang “Old and new tools to
dissect calcineurin's role in pressure-overload cardiac hypertrophy.” Cardiovasc
Res, vol. 53, no. 2, pp. 294-303, 2002. View at: Publisher Site | PubMed
[239] Beate Fiedler, Kai C Wollert
“Interference of antihypertrophic molecules and signaling pathways with the
Ca2+-calcineurin-NFAT cascade in cardiac myocytes.” Cardiovasc Res, vol.
63, no. 3, pp. 450-457, 2004. View at: Publisher Site | PubMed
[240] Olga G Shcherbakova, Carl M Hurt,
Yang Xiang, et al. “Organization of beta-adrenoceptor signaling compartments by
sympathetic innervation of cardiac myocytes.” J Cell Biol, vol. 176, no.
4, pp. 521-533, 2007. View at: Publisher
Site | PubMed
[241] Xin Li, Mohammed M Nooh, Suleiman W
Bahouth “Role of AKAP79/150 protein in beta1-adrenergic receptor trafficking
and signaling in mammalian cells.” J Biol Chem, vol. 288, no. 47, pp.
33797-33812, 2013. View at: Publisher
Site | PubMed
[242] Feng Zhu, Chi Yuan, Xu Zhang, et al.
“A-kinase anchoring protein 5-ancored calcineurin regulates the remodeling of
H9c2 cardiomyocytes exposed to hypoxia and reoxygenation.” Biomed
Pharmacother, vol. 155, pp. 113689, 2022. View at: Publisher Site | PubMed
[243] Madeline Nieves-Cintrón, Dinesh
Hirenallur-Shanthappa, Patrick J Nygren, et al. “AKAP150 participates in
calcineurin/NFAT activation during the down-regulation of voltage-gated K(+)
currents in ventricular myocytes following myocardial infarction.” Cell
Signal, vol. 28, no. 7, pp. 733-740, 2016. View at: Publisher Site | PubMed
[244] Chao Zeng, Jinyi Wang, Na Li, et al.
“AKAP150 mobilizes cPKC-dependent cardiac glucotoxicity.” Am J Physiol
Endocrinol Metab, vol. 307, no. 4, pp. E384-E397, 2014. View at: Publisher Site | PubMed
[245] Dario Diviani, Erica Reggi, Miroslav
Arambasic, et al. “Emerging roles of A-kinase anchoring proteins in
cardiovascular pathophysiology.” Biochim Biophys Acta, vol. 1863, no. 7
Pt B, pp. 1926-1936, 2016. View at: Publisher Site | PubMed
[246] V M Coghlan, B A Perrino, M Howard,
et al. “Association of Protein Kinase A and Protein phosphatase 2B with a
common anchoring protein.” Science, vol. 267, no. 5194, pp. 108-111, 1995.
View at: Publisher
Site | PubMed
[247] G A Jicha, C Weaver, E Lane, et al.
“cAMP-dependent protein kinase phosphorylations on tau in Alzheimer's disease.”
J Neurosci, vol. 19, no. 17, pp. 7486-7494, 1999. View at: Publisher
Site | PubMed
[248] Wang Cong, Xianglian Meng, Jin Li 1,
Qiushi Zhang, et al. “Genome-wide network-based pathway analysis of CSF
t-tau/Abeta1-42 ratio in the ADNI cohort.” BMC Genomics, vol. 18, no. 1,
pp. 421, 2017. View at: Publisher
Site | PubMed
[249] Nana Ma, Changrui Tie, Bo Yu, et al.
“Identifying lncRNA-miRNA-mRNA networks to investigate Alzheimer's disease
pathogenesis and therapy strategy.” Aging (Albany NY), vol. 12, no. 3,
pp. 2897-2920, 2020. View at: Publisher
Site | PubMed
[250] Luis F Santana, Manuel F Navedo
“Molecular and biophysical mechanisms of Ca2+ sparklets in smooth muscle.” J
Mol Cell Cardiol, vol. 47, no. 4, pp. 436-444, 2009. View at: Publisher Site | PubMed
[251] Manuel F Navedo, Edward P Cheng, Can
Yuan, et al. “Increased coupled gating of L-type Ca2+ channels during
hypertension and Timothy syndrome.” Circ Res, vol. 106, no. 4, pp.
748-756, 2010. View at: Publisher
Site | PubMed
[252] Pooneh Bagher, Christopher J Garland
“Scaffolding builds to reduce blood pressure.” Sci Signal, vol. 7, no.
333, pp. pe16, 2014. View at: Publisher Site | PubMed
[253] Jose Mercado, Rachael Baylie, Manuel
F Navedo, et al. “Local control of TRPV4 channels by AKAP150-targeted PKC in
arterial smooth muscle.” J Gen Physiol, vol. 143, no. 5, pp. 559-575,
2014. View at: Publisher
Site | PubMed
[254] Swapnil K Sonkusare, Thomas Dalsgaard,
Adrian D Bonev, et al. “AKAP150-dependent cooperative TRPV4 channel gating is
central to endothelium-dependent vasodilation and is disrupted in
hypertension.” Sci Signal, vol. 7, no. 333, pp. ra66, 2014. View at: Publisher Site | PubMed
[255] Sendoa Tajada, Claudia M Moreno,
Samantha O'Dwyer, et al. “Distance constraints on activation of TRPV4 channels
by AKAP150-bound PKCalpha in arterial myocytes.” J Gen Physiol, vol.
149, no. 6, pp. 639-659, 2017. View at: Publisher Site | PubMed
[256] Zishao Zhong, Zhenhao Ye 1 2, Guihua He, et al. “Low expression of A-kinase anchor protein 5 predicts poor prognosis in non-mucin producing stomach adenocarcinoma based on TCGA data.” Ann Transl Med, vol. 8, no. 4, pp. 115, 2020. View at: Publisher Site | PubMed
[257] Una Kjällquist, Rikard Erlandsson, Nicholas P Tobin, et al. “Exome sequencing of primary breast cancers with paired metastatic lesions reveals metastasis-enriched mutations in the A-kinase anchoring protein family (AKAPs).” BMC Cancer, vol. 18, no. 1, pp. 174, 2018. View at: Publisher Site | PubMed