Received: Tue 23, May 2023
Accepted: Mon 12, Jun 2023
Abstract
Breathing is a natural process that we cannot do without and is also a daily action that sensitively and intensely changes under various situations. What if this essential act of breathing can impact our overall well-being? Recent studies have demonstrated that breathing couples with higher brain functions, i.e., perception, motor actions, and cognition. In particular, the timing of breathing can be a key factor to modulate accuracy in cognitive tasks and changes activation in specific cortical regions. To determine possible respiratory roles in attentional and memory processes and functional brain networks, we discussed several issues of interactions between breathing and brain function: i) respiration-dependent modulation of cognition and mental health; ii) respiratory rhythm generation in the brainstem (e.g., the PreBötzinger complex); iii) interpretation of the blood-oxygen-level-dependent (BOLD) signal without respiratory artifacts using functional magnetic resonance imaging (fMRI); iv) respiration-timing-dependent effects on functional neural networks (e.g., the locus coeruleus, temporoparietal junction, ventral attention network, and cingulo-opercular salience network); and v) a potential application of breathing manipulation in mental health care. These outlines and considerations of “brain-breath” interactions lead to a better understanding of the interoceptive and cognitive mechanisms that underlie brain-body interactions in health conditions and in stress-related and neuropsychiatric disorders.
Keywords
Inspiration, expiration, supramarginal gyrus, inferior parietal lobule, middle frontal gyrus, dorsomedial prefrontal cortex, anterior cingulate cortex, anterior insula, interoception, encoding, retrieval, recognition, stress
1. Introduction
We have control over our own bodies and can perform actions voluntarily as planned and expected. For example, as reading this sentence, we are moving our eyes across the text and wondering about what we are about to read next. Such regulatory mechanisms of brain function are defined in a “top-down” fashion. On the other hand, we hear our own heartbeats racing when we stand in front of a large audience for playing a baseball or making a symposium presentation. Such heartbeats are involuntarily regulated according to autonomic mechanisms based on the roles of homeostasis and allostasis, and are defined as vegetative function in a “bottom-up” fashion.
Pertinently, there are interoceptive pathways that influence brain function [1]. Regarding the concept of the brain as a prediction machine (i.e., the free-energy principle; [2, 3]), it has been hypothesized that when afferent interoceptive inputs are noisy or imprecise at least in the short term, top-down predictions reduce attention to mismatch or ignore bottom-up inputs, which convey prediction errors [4]. This misinterpretation may cause prolonged or inadequate responses of cellular and molecular mechanisms, resulting in allostatic load, cognitive deficits, and stress disorders [5, 6]. It remains unclear whether top-down regulation can precisely coordinate bottom-up regulation.
Breathing is a natural process that we cannot do without; however, it is also said that breathing is a daily action that sensitively and intensely changes under various situations. The respiratory control system is distinct from other autonomic systems and is unique because it has innervation with top-down regulation of consciousness [7]. Recent studies have provided evidence on how the electrophysiological activity of the brain, sensation, motor actions and cognition depend on the phase of the respiratory cycle (Figure 1) [8-11]. Oku described that the breathing process has complex variability that is caused by the respiratory central pattern generator (CPG) and discussed obscure information that can be translated into a form in clinical practice for diagnosis, emotion, and mental conditioning. We recently found that breathing changes the accuracy of cognitive tasks; i.e., the accuracy of a delayed matching-to-sample (DMTS) task was decreased when the retrieval process spanned the exhalation-to-inhalation (EI) transition during the respiratory cycle [12, 13].
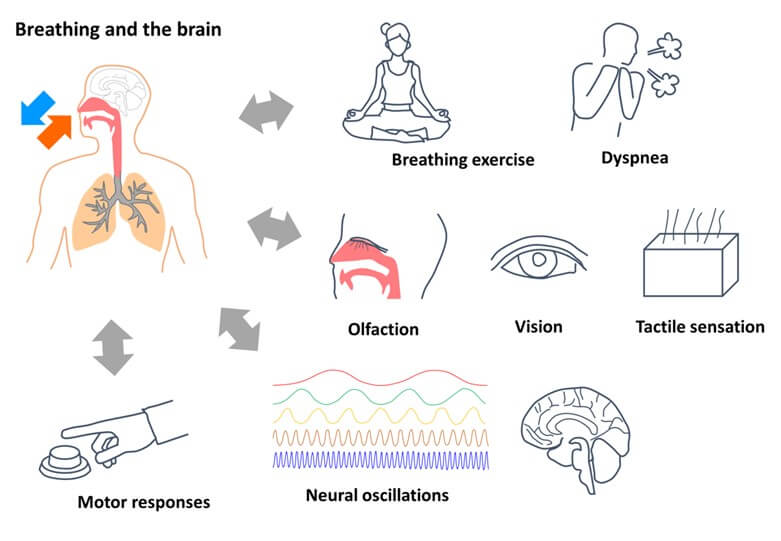
Breathing and the brain have crosstalk interactions. Brain states are changed by breathing exercises and dyspnea. Respiration couples with olfaction, vision, tactile sensation, and cortical neural oscillations. Furthermore, successful motor responses in cognitive tasks are associated with certain times during the respiratory cycle.
Notably, the EI transition-dependent effects were relevant to activation in specific cortical regions, specifically the temporoparietal junction (TPJ, also known as the supramarginal gyrus and inferior parietal lobule or IPL), which is a neural core of the ventral attention network (VAN) and has preferential functional connectivity to the cingulo-opercular network (CON) [14, 15]. Related to these findings, we review several issues of interactions between breathing and brain function. First, we introduce phenomena, in which breathing couples with attention, sensation, motor actions, and cognition, and affects mental health. Second, we explain neural mechanisms generating respiratory activity and phase components (e.g., the EI transition) in the brainstem. Third, we introduce the primary human neuroimaging method of functional magnetic resonance imaging (fMRI), which can cause respiratory artifacts in blood-oxygen-level-dependent (BOLD) signals, and then describe how to interpret BOLD signals without respiratory artifacts. Finally, we discuss the potential roles of respiration that spontaneously changes large-scale brain network activity and how it could be applied as a possible mental health treatment to reduce or eliminate symptoms of stress-related and neuropsychiatric disorders.
2. Respiration and Brain States
2.1. Attention to Breathing
Historically, the act of breathing has long been believed to shape mind and affect mental conditions. Breathing exercises, such as slow-deep breathing and paying attention to each breath, modify cardiovascular and brain functions and consequently improve mental health and cognitive and motor performance [16-19]. Breathing practices are typically incorporated in training methods for Zen meditation and yoga, such as transcendental meditation, 4-7-8 breathing, and Sudarshan kriya yoga, and are used to alleviate psychiatric and stress-related medical conditions [20-21]. There is substantial evidence that breathing contributes to improvements in brain states. Human neuroimaging studies showed that spontaneous breathing was coordinated with periodic brain activity during resting states, which were measured by fMRI [22] and magnetoencephalography (MEG) [23]. Other studies using intracranial electroencephalography (iEEG) and fMRI indicated that when healthy volunteers paid attention to their breathing, respiration-brain signal coupling (i.e., respiration-iEEG coherence and respiration-fMRI signal synchronization) was increased in the insula, anterior cingulate cortex (ACC), premotor cortex, and hippocampus [24, 25]. Respiration-iEEG coherence also increased in the frontotemporal-insular network when healthy participants breathed at a voluntary and slightly faster rate [24]. Respiratory sensation is likely to involve the insula, which is known as a central hub for interoception [26-28].
2.2. Dyspnea
Dyspnea is a tightening feeling of not being able to breathe and take in enough air, and it is distinguished from the breathlessness we would observe in normal subjects, such as that induced by exercise. Dyspnea is defined as a subjective experience of breathing discomfort that consists of qualitatively distinct sensations that vary in intensity [29-31]. Notably, a human lesion study demonstrated that the insula was associated with sensitivity to dyspnea and pain; patients with lesions of the right insula showed reduced sensitivity in the perception of unpleasant sensations, particularly in the perception of dyspnea and pain [32]. Dyspnea and palpitations can be compulsorily caused by isoproterenol, an agonist of peripheral beta-adrenergic receptors. Hassanpour et al. [32] found that the injection of isoproterenol led healthy volunteers to exhibit increased dyspnea and palpitations with concomitant fMRI activation of the right insula. During the recovery process, fMRI activation of the right insula spread from the right anterior insula to the right posterior insula and to the left middle insula. Von Leupoldt et al. [34] revealed that respiratory load can induce activation of the insula. Negative emotional stimulation (i.e., viewing a standardized emotional picture series from the international affective picture system or IAPS Database [35]) successfully induced respiratory load, which was associated with higher levels of unpleasantness of perceived dyspnea when compared with respiratory load during positive emotional stimulation. Then, a negative emotional-stimulated respiratory load increased fMRI activation in the right anterior insula and right amygdala, suggesting that dyspnea-related unpleasantness is processed in the right anterior insula and amygdala [34]. Accordingly, these findings indicate that the right anterior insula and its associated regions may play a crucial role in respiratory sensation and dyspnea.
In terms of medical psychiatric phenomena, chronic obstructive pulmonary disease (COPD) is a respiratory disease that chronically causes airflow blockage and breathing-related problems and is associated with a high incidence of anxiety and panic attack symptoms [36, 37]. The breathing pattern is exaggerated in excess of metabolic need and can cause dyspnea and, as a result, bring on panic attacks in patients [38]. These patients can misinterpret normal bodily sensations and become almost obsessively focused on something being wrong [36]. It is suggested that this misinterpretation might be associated with dysfunction or overactivity of the anterior insula that is implicated in anxiety disorders characterized by burdensome preoccupation with somatic symptoms [39].
2.3. Respiration and Sensation
It is said that breathing contributes to the detection of sensory information from the outside world. Rodent studies have shown that sniffing or sampling of olfactory information is coordinated during the respiratory cycle [40]. In awake rats, activity of the mitral and tufted cells in the olfactory bulb was coupled to certain times of inhalation, resulting in successful odor discrimination [41, 42]. Whisking (vibrissa touch) detection was phase-locked to certain times of exhalation, in coordination with a phase shift of sniffing to compensate for output from the olfactory bulb [43, 44]. Human studies have identified a similar tendency in respiratory phase locking to sensation, i.e., olfaction, touch, and vision. Odor detection occurring during inhalation was phase-locked to alpha oscillations, but it was not seen in the inspiration-triggered potentials of normal breathing [45]. Respiratory phase locking to tactile sensation showed the highest detection rate during the early phase of exhalation [46]. Visual signals presented during exhalation were detected more frequently than those presented during inhalation [47]. A recent study using MEG found that the sampling of visual information was preferentially aligned with cortical activity at a certain point in the respiratory phases to facilitate visual sensitivity [48]. Notably, maximized cortical activity occurred during the late phase of inhalation. Although respiratory phase locking to sensory detection was occasionally shifted at certain times during the respiratory cycle, respiration may play an important role in enhancing discrimination rates of sensory information.
2.4. Respiration and Neural Oscillations
Numerous studies using laboratory rodents have determined coupling between breathing and brain oscillatory activity [49, 50], the latter of which was measured by local field potential (LFP) and electroencephalography (EEG). In rodents, the respiratory rhythm entrains delta (0.5-4 Hz), theta (4-12 Hz), and gamma (30-80 Hz) neural oscillatory ranges in the olfactory bulb and extensive cortical regions, i.e., the piriform cortex [51], barrel cortex [52], hippocampus [53-56], and prefrontal cortex [57, 58]. Respiration-coupled oscillations, which were identified in mice in awake states [52, 53], were slow in their synchronized frequency (i.e., delta oscillations) during immobility and were faster (i.e., theta oscillations) during exploratory behavior [57, 58]. Respiration-delta/theta coupling in rodents was abolished by breathing after olfactory bulbectomy [52, 57] or with direct trachea airflow instead of nasal airflow (tracheotomy) [55, 56], suggesting that the airflow in the respiratory system through the nasal cavity may be essential for the formation of respiration-delta/theta coupling. Although the respiratory frequency is determined by the body size of mammals during awake states (i.e., 3-7 Hz in mice, 1-3 Hz in rats, and 0.15-0.3 Hz in humans), respiration-locked activity occurs in the human brain, i.e., the piriform cortex and hippocampus in epilepsy patients as determined using iEEG recordings [24, 59]. In an extensive analysis of human data, the phase-amplitude coupling between respiration (i.e., 0.15-0.3 Hz) and hippocampal theta was detected at 0.28 Hz [50]. Moreover, while respiration coupled with gamma oscillations in the hippocampus and prefrontal cortex [56-58], theta-gamma coupling in the hippocampus and prefrontal cortex is associated with successful cognitive performance in rodents [60-62] and humans [63-65]. Importantly, the disappearance of respiration-delta/theta coupling diminished gamma oscillations in mice [52]. Hence, respiration-delta/theta coupling might be crucial for the generation of gamma oscillations during cognitive brain states [50].
2.5. Respiration and Motor Actions
Studies of animal ecology have revealed that respiration is phase-locked to locomotion in wild mammals. For instance, a wallaby runs across the prairie field while hopping with both feet together. The ratio of breathing to hopping is phase-locked at a 1:1 ratio, whereas the heart rate is not synchronized with hopping [66]. In bats, the ratio of breathing to wing flapping is synchronized at a 1:1 ratio [67]. A dog coordinates diaphragmatic oscillations and chest wall deformation during trotting behavior [68]. In humans, numerous studies have shown that the respiratory phase reflects walking, running, and successful motor actions. When healthy volunteers started manipulating an object with their fingers, performance accuracy was dependent on coordination in either inhalation or exhalation with internal representations during either adaptative or predictive processes [69, 70]. Notably, Park et al. [71] demonstrated that voluntary motor actions were more preferentially initiated during the late phase of exhalation, and one-at-a-time cortical activity was coordinated with motor actions. In healthy volunteers performing memory tasks and discrimination tasks, button-press responses by fingers occurred more frequently during exhalation [72] and the late phase of exhalation [12]. However, simple, externally triggered actions did not show respiratory phase locking [71]. Thus, it is likely that successful motor responses in cognitive tasks are associated with certain times during exhalation.
2.6. Respiration and Cognitive Processes
Considerable evidence has been provided on how respiration modulates cognitive processes and brain function. Cognitive processes are a mixture of subprocesses during online brain states, e.g., attention, encoding, retrieval, and decision-making, as well as offline brain states, e.g., sleep and consolidation. Human studies have shown that nasal respiration during encoding, consolidation, and retrieval of recognition memory tasks increased accuracy compared to oral respiration [59, 73]. However, phase-dependent effects of respiration (e.g., inhalation vs. exhalation) on accuracy are more complicated [59, 72]. In an eyeblink conditioning task in healthy volunteers, the rate of conditioned responses was higher when they were trained at exhalation compared to inhalation [74]. Zelano et al. [59] found that accuracy was higher when test cues (i.e., retrieval cues) were presented during inhalation than exhalation in a recognition memory task. It should be noted that retrieval and recognition processes have an array of cognitive subprocess components, e.g., accessing what-where-when information, components of accessibility and availability, and match and mismatch processes [75-77]. Thus, a key component must be whether the retrieval process crosses phase transitions during the respiratory cycle, i.e., the EI transition (or the onset of inhalation) and inhalation-to-exhalation (IE) transition (or the onset of exhalation), at which different sets of respiratory neurons in the brainstem start firing synchronously [78, 79]. Indeed, Perl et al. [80] showed that accuracy was increased when test cues were presented at the exact time of EI transition (onset of inhalation) than at the exact time of IE transition (onset of exhalation) in a visuospatial task (or mental-rotation task).
We have provided evidence that phase transitions (i.e., EI transition vs. IE transition) differentially affect accuracy and reaction time (RT) when the retrieval process spans each transition during the DMTS recognition memory task with a short delay [12, 13]. RT is a popular measure of cognitive function and refers to the duration between the presentation of a sensory cue (e.g., light or sound) and a button-press response in cognitive tasks. RT is a critical indicator of discrimination ability and reflects task difficulty. Our findings showed that accuracy was reduced and RT was extended when the retrieval process spanned the EI transition compared to other respiratory components (e.g., the IE transition), suggesting that the EI transition must be a key component for modulating cognitive performance and brain function. We propose that the EI transition might play critical roles in both a “reset and trigger” during cognitive processes, possibly leading to increased accuracy when the EI transition starts at the onset of the process [80] and decreased accuracy when the EI transition occurs in the middle of the process [12, 13]. Next, we discuss the neural mechanisms generating the EI and IE transitions during respiratory cycles.
3. Neural Mechanisms of Respiratory Rhythm Generation
3.1. Respiratory Central Pattern Generator (CPG)
It has been suggested that respiratory CPGs determine breathing activity, which alternates between inhalation and exhalation with combinations of tidal pressure (amplitude) and phase duration (time), although the complete picture of the respiratory CPG remains unclear. The primary inspiratory rhythm generator was found in the PreBötzinger complex (PreBötC), which is a bilateral neuronal nucleus in the ventrolateral medulla [81]. On the other hand, the expiratory rhythm generator was identified in the parafacial respiratory group (pFRG, also known as the lateral parafacial nucleus [82]), which is bilaterally located ventrolateral to the facial nucleus in the medulla and partially overlaps with the retrotrapezoid nucleus. However, the pFRG appears to be quiescent in mature intact rodents and generates late-expiratory bursts conditionally [83, 84]. Therefore, the PreBötC is considered the kernel for respiratory rhythm generation [81, 85].
3.2. A Neural Mechanism of the Exhalation-to-Inhalation (EI) Transition
A neural mechanism that generates the EI transition is different anatomically and physiologically from the mechanism that generates the IE transition (Figures 2 & 3). The respiratory cycle, meaning alternation between inhalation and exhalation, is formed by dynamic interactions between excitatory and inhibitory neurons of the respiratory CPG in the medulla and pons (Figure 2a) [78, 79, 86, 87]. The respiratory cycle consists of three distinct phases (Figure 2b): inspiration (inhalation), postinspiration (the early phase of exhalation), and active expiration (E2, the late phase of exhalation). Hence, the EI transition and IE transition are defined by the onsets of inspiration and postinspiration, respectively. “Medullary” respiratory oscillators, causing a biphasic inspiratory-expiratory rhythm, are composed of networks between inspiratory excitatory neurons (i.e., pre-I/I neurons, Figure 2a) in the PreBötC and the “inhibitory ring” of three types of inhibitory neurons (i.e., early-I, post-I, and aug-E neurons) in the PreBötC and adjacent Bötzinger complex [78]. During inspiration, the subpopulation of inspiratory excitatory neurons (pre-I/I neurons), which are bilaterally connected to excitatory glutaminergic neurons [88-89], persistently bursts intrinsically even when the PreBötC is isolated in vitro in a slice from a neonatal rodent medulla (Figures 3a & 3b) [81, 90]. As demonstrated, according to the property of excitatory neurons in the PreBötC, the EI transition can be generated in an “abrupt and divergent manner” (Figure 3a).
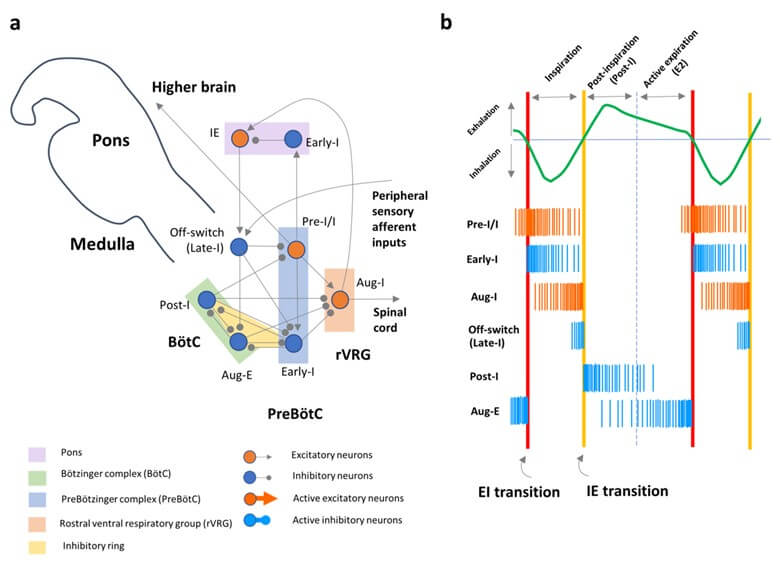
a) Schematic drawing showing network connectivity with subtypes of respiratory neurons in the medulla and pons [78, 79, 86]. b) Drawing showing firing patterns of individual respiratory neurons in the medulla during the respiratory cycle [78, 79]. A respiratory waveform, a green line; the exhalation-to-inhalation (EI) transition, red lines; the inhalation-to-exhalation (IE) transitions, yellow lines; excitatory neurons, bundles of orange lines; and inhibitory neurons, bundles of blue lines.
3.3. A Neural Mechanism of the Inhalation-to-Exhalation (IE) Transition
A resetting of oscillatory processes provides gate control of neuronal excitability that irreversibly terminates inspiration and blocks afferent excitatory synaptic input from the network and periphery and then starts the IE transition and subsequent postinspiration (Figures 3c-3e) [78]. Numerous studies suggest that the postinspiration phase can be caused by an inspiratory off-switch mechanism via i) the sensory feedback from the inflated lung arising from the pulmonary stretch receptor, known as the Breuer-Hering reflex [91], and ii) intrinsic synaptic mechanisms in the pons [86]. In particular, the inspiratory off-switch is initiated by phasic or tonic firing neurons (i.e., IE neurons) in the Kölliker-Fuse (KF) nucleus and adjacent subnuclei of the parabrachial complex in the pons (Figure 3c) [78, 92]. Then, postinspiratory inspiratory neurons (i.e., Post-I neurons) in the Bötzinger complex fire to prevent the lungs from abrupt deflation during the postinspiratory phase (Figure 3d).
Anderson et al. [93] also demonstrated that excitatory neurons in the intermediate reticular nucleus in the medulla function as independent oscillators of the postinspiratory complex (PiCo) with rhythm-generating but distinct modulatory properties. As a result, it is believed that the PiCo might independently contribute to the generation of postinspiratory behaviors, such as swallowing and vocalization [87, 94]. However, the blockade of these excitatory activities by the GABAA receptor agonist isoguvacine did not cause prolongation of inspiration or apneusis [95]. Thus, the hypothesis of the PiCo as an independent oscillator remains under debate.
Moreover, bilateral lesions of the KF in the pons or vagotomy (for the Breuer-Heiring reflex) cause severely prolonged inspiration or apneusis and can destroy the timing of the inspiratory off-switch [86, 96]. These findings indicated that either KF input or vagal afferent input, or both, act through the inspiratory off-switch mechanism involved in the proper timing of IE transition. Nevertheless, apneusis after KF lesions and vagotomy was not permanent because networks outside the sensory-pontine loop compensate for the lack of the function of the inspiratory off-switch and perhaps because afferent inputs from cortical regions (i.e., ACC) control conscious and behavioral adjustment of breathing [7, 97]. As a result of an irreversible termination setup from a diversity of neural networks within and outside the medulla and pons, the IE transition is implied to be generated in a “convergent manner” (Figures 3c & 3d).
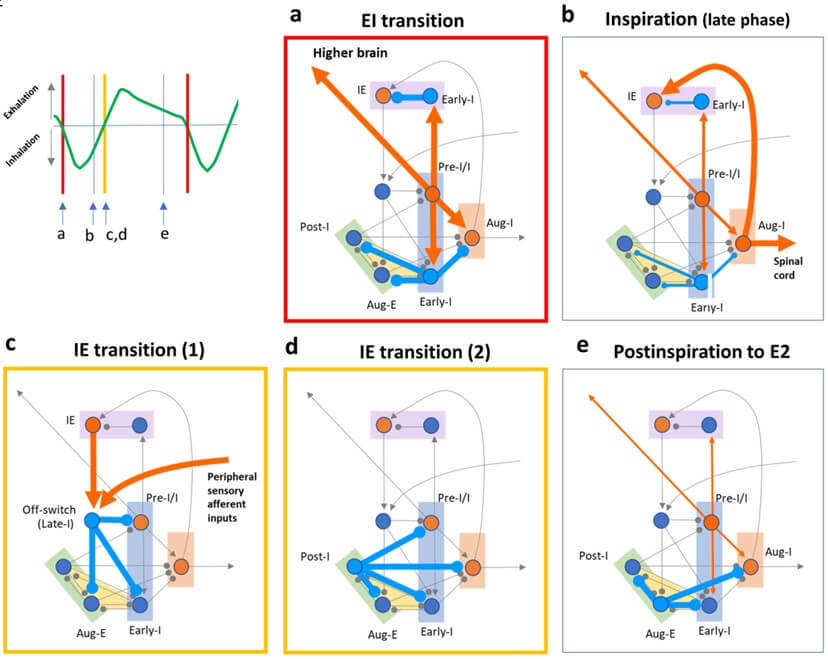
Drawing showing combinations of firing patterns of respiratory neurons at each phase during the respiratory cycle (left upper panel). a) At the EI transition (a red rectangle), excitatory Pre-I/I neurons (orange) in the PreBötzinger complex (PreBötC) activate inhibitory Early-I neurons (blue) in the PreBötC and pons. b) Excitatory Aug-I neurons (orange) in the rostral ventral respiratory group (rVRG) fire during the late phase of inspiration. c & d) At the IE transition (yellow rectangles), excitatory IE neurons (orange) in the pons and peripheral sensory afferent inputs activate inhibitory off-switch neurons (Late-I neurons, blue) in the medulla, and then inhibitory Post-I neurons (blue) fire in the BötC. e) Inhibitory Aug-E neurons (blue) fire in the BötC during postinspiration and active expiration (E2).
4. Respiration and fMRI
4.1. BOLD Signal Measured by fMRI
To date, respiration-dependent effects on the human brain have been identified using neuroimaging approaches characterized by different temporal and spatial resolutions, such as noninvasive EEG [45, 80], intracranial EEG [24, 59], MEG [23, 48], positron emission tomography (PET), and fMRI. An early PET study showed that active inhalation and active exhalation induced activation in the primary motor cortex and supplementary motor area (SMA) [98]. There is a growing consensus that fMRI is the primary neuroimaging method in human research that can determine task-related activation and resting-state functional connectivity. However, since fMRI signals sensitively reflect cerebral CO2 fluctuations and breathing patterns, fMRI data targeting respiration-dependent effects must be processed carefully so that the data are appropriately interpreted as neural activity [99].
The fMRI signal, which is known as BOLD contrast [100], is a complex hemodynamic response function (HRF) underlying metabolism (i.e., cerebral metabolic rate of oxygen consumption or CMRO2), vasodilation (i.e., cerebral blood volume or CBV), and perfusion (i.e., cerebral blood flow or CBF) in the cerebral cortex (Figure 4a) [101, 102]. Meanwhile, neural activity can be classified by LFP and spiking activity; LFP represents extracellular input into neurons, while spiking activity from cell bodies with a suprathreshold represents intracellular output. Although the relationship between BOLD signals and neural activity remains a matter of debate, it has been shown that BOLD signals are correlated with i) LFP rather than spiking activity [103, 104]; ii) mostly spiking activity; and iii) both LFP and spiking activity [101]. Increased neural activity is accompanied by an increased flux of Na+, K+, and Ca2+ ions and increased ATP production via glucose consumption. These changes may induce a coupling between neurons and astrocytes to send vasoactive signals into nearby arterioles and capillaries, consequently dilating the upstream arteries (i.e., neurovascular coupling; Figure 4a) [105]. Then, BOLD signals can be an indicator of synchronous neural activity [101].
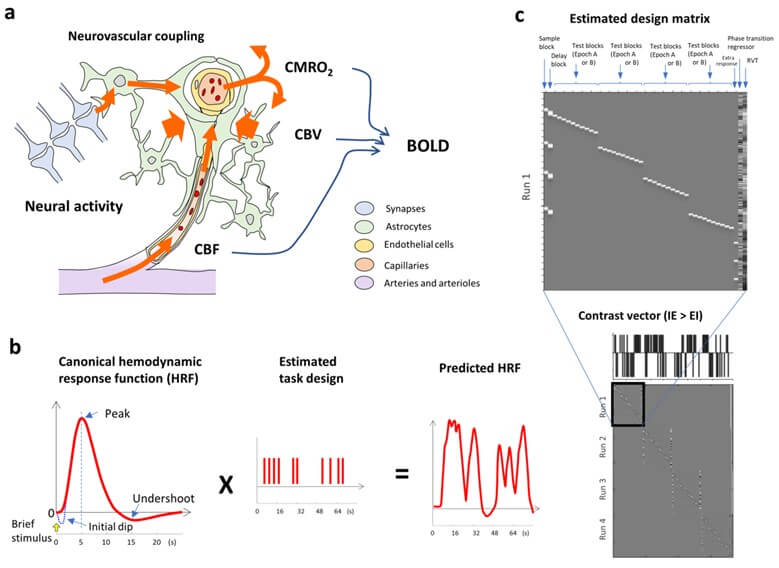
a) Changes in BOLD signals are determined by three parameters: i) the cerebral metabolic rate of oxygen consumption (CMRO2), ii) cerebral blood volume (CBV), and iii) cerebral blood flow (CBF) in the cerebral cortex. Thus, BOLD signals are an indirect parameter of neural activity through neurovascular coupling mechanisms [102, 105]. b) Generally, BOLD-related neural activity is estimated by a neural model convolved with the canonical hemodynamic response function (HRF) using the general linear model. A time series of BOLD-related neural activity is detected as the predicted HRF that was calculated by the canonical HRF and a time series of an estimated task design. c) Matrix showing an example of an estimated design matrix containing 40 task blocks during a single fMRI run (upper panel) and an estimated design matrix at an individual level (four fMRI runs, lower panel) with the contrast vector between the IE and EI conditions (i.e., IE > EI) according to single participant performance [12].
4.2. Respiratory Artifacts in BOLD Signals
Changes in BOLD signals accurately reflect neural activity if the intermediate vascular steps are not significantly altered [102]. Arterial CO2 concentration alters with respiratory depth and rate, leading to BOLD signal changes that are unrelated to neural activity. Such fluctuations in arterial CO2 concentration cause variations in respiration volume per time (RVT) [106, 107]. In particular, deep breathing and breath holding strongly contribute to BOLD signals, which reflects a severe respiratory artifact [99, 108]. Huijbers et al. [108] showed that retrieving visual items that were encoded during a 20-s period of breath hold decreased fMRI activation in the posterior midline brain region compared with retrieving items that were encoded during spontaneous breathing [108]. Meanwhile, no difference in fMRI activation was observed in the ventrolateral prefrontal cortex between breath holding and spontaneous breathing conditions. Notably, the effects of deep breath, breath hold, and RVT can be mostly regressed out using the AFNI program of the RetroTS algorithm [106, 109, 110].
Even though respiratory artifacts are mostly removed from BOLD signals, neural activity derived from the PreBötC is problematic to be interpreted as BOLD-related neural activity. A time series of BOLD-related neural activity is usually detected by a neural model convolved with canonical HRF using the general linear model (Figures 4b & 4c) [111, 112]. However, BOLD signals contain noise and anatomical and physiological artifacts that are unrelated to neural activity. Generally, fMRI data are processed through the following multiple steps before data analysis: i) corrections for image distortions induced by magnetic field nonlinearity and static magnetic field susceptibility; ii) nonlinear image normalization to Montreal Neurological Institute (MNI) space; iii) intensity normalization; and iv) denoising, such as by independent component analysis (ICA)-based denoising of time series data using multirun application of FMRIB’s ICA-based Xnoiseifier (multirun ICA-FIX) [12, 113, 114]. Recently, the multirun ICA-FIX approach has gained attention: a time series of fMRI data was divided into spatial map, temporal map, and power spectrum components and classified as either signal or noise components semiautomatically based on certain criteria [113, 115]. In particular, the cardiac and respiratory cycles, which appear as low-frequency fluctuations at approximately 1 Hz and 0.3 Hz, respectively, can be extracted as components of predominant powers. Thus, neural activity derived from the respiratory rhythm generator the PreBötC may be difficult to detect as BOLD-related neural activity because a series of mechanical and physiological respiratory functions always have the same or concomitant rate and patterns for respiratory neuron-based activity. Based on the denoising approach, it is reasonable to mention that the EI-transition-dependent fMRI signal might not reflect rhythmic PreBötC activity itself; instead, this signal represents neural activity that is accompanied by PreBötC activity or derived from PreBötC-associated neural activity during the task.
5. Respiration and Functional Neural Networks
5.1. Respiration-Dependent Signaling Pathways to the Cerebral Cortex
Our findings have thus far shown that the decline in cognitive performance was specific to the EI transition during the retrieval process, while an extensive array of frontoparietal regions were activated during the encoding, delay, and retrieval processes in the DMTS task (Figure 5) [12]. When the retrieval process spanned the EI transition (EI condition), activation was reduced in the anterior cluster of the right temporoparietal junction (TPJa), right middle frontal gyrus (MFG), and left and right dorsomedial prefrontal cortex (dmPFC, containing the dorsal part of the ACC) compared to other respiratory conditions during the retrieval process (i.e., IE condition, INS condition, and EXP condition; (Figure 5b), (left panel in Figure 5d). However, activation in these regions during the EI condition was still higher than activation during the encoding process or the delay period of the task (right panel in Figure 5d). Our results suggested that the coordination between the timing of respiration and specific cortical networks could be a key driver in modulating brain function, thereby influencing subsequent task performance [12].
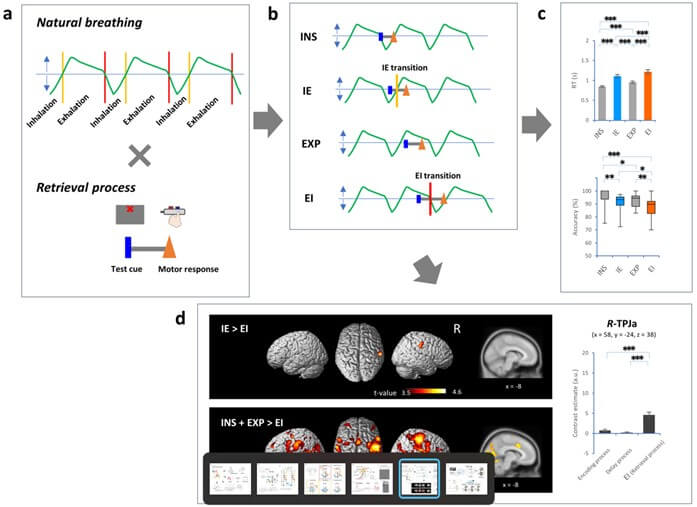
a) In healthy volunteers performing the DMTS task, the retrieval process occurred during natural breathing. The retrieval process was defined by the period from a test cue to a motor response. b) There were four conditions of crossing between natural breathing and the retrieval process: i) the retrieval process fitting within inhalation (INS condition), ii) the retrieval process encompassing the IE transition (IE condition), iii) the retrieval process fitting within exhalation (EXP condition), and iv) the retrieval process encompassing the EI transition (EI condition). c) Plots showing that the EI condition exhibited the longest RT and lowest accuracy in the respiratory conditions. d) Images showing brain regions that exhibited fMRI activity in the IE condition in contrast to the EI condition (IE > EI, left upper panel). Images showing brain regions that exhibited fMRI activity in the INS + EXP condition in contrast to the EI condition (INS + EXP > EI, left lower panel). Plots showing a contrast estimate in the right TPJa (MNI coordinates: x = 58, y = -24, z = 38 [12]). fMRI activity was exhibited in the region of interest at an SPM{t} threshold of p < 0.05 with cluster-level FWE correction for the whole brain.
a.u.: arbitrary unit.
Here, we address the question of whether the signal derived from PreBötC activity or PreBötC-associated activity is transmitted to the TPJa, MFG, and dmPFC mediated by specific neural hubs. Recent studies have suggested candidate neural hubs for the olfactory bulb [50], thalamus [116], parabrachial nucleus [117], and locus coeruleus (LC) [118]. Our findings suggest that respiratory modulation could be relevant to attention mechanisms based on the pathway mediated by the LC during retrieval [118-120], even though other neural hubs cannot be ruled out. The LC is implicated in attention and arousal [122, 123] and has two modes of activity [119]: phasic-mode activity that corresponds to task engagement (or exploitation) with high performance and enhances cortical activity of salience information [124] and tonic-mode activity that is associated with the absence of phasic activity and task disengagement (or exploration). Next, we consider the possibility of TPJa and LC function originating from PreBötC activity or PreBötC-associated activity.
5.2. Inferior Parietal Lobule (IPL)/Temporoparietal Junction (TPJ)
We found that the EI transition had primarily reduced fMRI activation in the region that lies anatomically along the boundary between the IPL and TPJ. According to Mars et al. [125] and Igelstrom et al. [126], this region is classified as the right TPJa (MNI coordinates: x =58, y = -24, z = 38). The right TPJa is well known as a neural core of the VAN, together with the right MFG as another neural core, which is a bottom-up system involved in attention and awareness of surprising or salient events from the external environment [14, 121, 127]. The right MFG might be linked to the dorsal attention network, which may be involved in top-down attentional control. Furthermore, resting-state fMRI studies have shown that the right TPJa has preferential functional connectivity to the CON (also known as the salience network or SN), whose neural hubs are the dorsal part of the ACC and anterior insula, which may be involved in alertness and salience detection [15, 128]. These functional connectivity studies support the idea that the VAN and CON/SN serve as macroscopic anatomical substrates to cross-link functional networks and share information (Figure 6a) [15, 129-131].
Human studies have also suggested that the TPJ is one of the most prominent sources of P300 (P3) components, which are event-related potentials with a latency of 300-400 ms [120]. P300 components are associated with surprising events [132] and are hypothesized to reflect the updating of information in anticipation of subsequent information processing [120, 133]. Chiefly, P300 components are assumed to reflect the phasic-mode activity of the LC [119, 134]. One human pharmacological study showed that while oddball target responses induced P300 components in the left and right TPJ and prefrontal cortex, these P300 components were abolished by an injection of the beta-adrenergic blocker propranolol [135]. Interestingly, a resting-state fMRI study revealed LC-to-right TPJ functional connectivity [136]. Liebe et al. [137] demonstrated that patients with mild cognitive impairment (MCI) had reduced LC-to-right TPJ functional connectivity but elevated LC-to-ACC connectivity and LC-to-left anterior insula connectivity. Moreover, LC-to-right TPJ connectivity was positively correlated with memory scores in dementia patients, supporting our findings that showed a positive correlation at the within-individual level between right TPJa activation and the discriminability score for memory (Figure 6b) [12]. Liebe et al. [137] suggested that an increase in LC-to-ACC, LC-to-anterior insula, and SN connectivity might result in elevated tonic-mode activity of the LC [119], whereas a decrease in LC-to-right TPJ connectivity could be correlated with decreased phasic-mode activity of the LC [14].
5.3. PreBötzinger Complex (PreBötC) and Locus Coeruleus (LC)
The possibility that signals from the PreBötC modulate the LC was suggested by genome-wide association and molecular biological approaches. Yackle et al. [118] showed that Cdh9/Dbx1 double-expressing PreBötC neurons had efferent projections specifically to the LC. Then, conditional, bilateral genetic ablation of Cdh9/Dbx1-expressing PreBötC neurons in adult mice left breathing intact but increased calm behavior and decreased time in arousal states, suggesting that the PreBötC regulates noradrenergic neurons in the LC [118].
Although it remains unclear whether PreBötC-to-LC-dependent effects cause memory decline and deactivation of the right TPJa, studying patients with posttraumatic stress disorder (PTSD) may provide insights into the functional relationship of LC activity during cognitive processes [138]. Normally, LC activity slows down during the state of rapid eye movement (REM) sleep and is also silent in the seconds immediately preceding sleep spindles during non-REM sleep. Furthermore, the characteristic sleep disturbance of PTSD, insomnia, and opiate withdrawal suggest overactivity of the LC during sleep, which may underlie emotional and hippocampal memory consolidation deficits in patients with these disorders [122]. Importantly, PTSD patients have increased heart rates, skin conductance, eye blink, and fMRI activation in the LC and right intraparietal sulcus in response to loud sounds compared with trauma-exposed controls [139]. These ideas propose that decreasing LC activity may be relevant to resetting the novelty of information processing and then immediately starting consolidation (and reconsolidation).
5.4. Roles of the Ventral Attention Network (VAN)/Cingulo-Opercular Network (CON)/Salience Network (SN)
At this stage, the role of the EI transition that reduced activation of the VAN/CON/SN and caused cognitive decline and decreased accuracy of the task remains puzzling (Figure 6a) [12]. However, recent findings have given significant consideration to interactions between mental health problems and functional neural networks. The SN is known as a network hub that may receive interoceptive information for an adaptive response, wherein the SN disengages the default mode network and engages the lateral frontoparietal (or central executive) network in mediating attention, working memory, and other cognitive processes for goal-oriented behavior [128, 140, 141]. Salience processing provides a motivational context for a stimulus and may thus be central to initiating motivated or resetting behavior. Conversely, aberrant salience processing may underlie motivational disturbances [142]. In humans, dysfunction and/or hyperactivity of the SN are implicated in psychiatric disorders [143, 144], such as depression [142, 145], schizophrenia [146, 147], PTSD [148, 149], and traumatic brain injury [150, 151]. Therefore, we propose that normalization of respiratory properties using breathing manipulation, e.g., rhythmic inhalation and exhalation in synchronization with sensory detection and motor responses, could be a target in mental health care to reduce or eliminate symptoms in patients with stress responses and neuropsychiatric disorders.
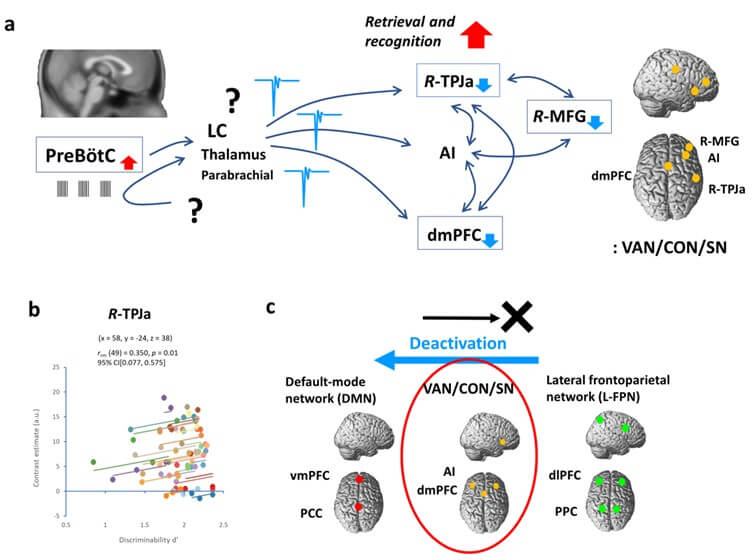
a) Drawing showing EI-transition-dependent modulation of the retrieval and recognition processes through a potential pathway from the PreBötC to the ventral attention network/cingulo-opercular network/salience network (VAN/CON/SN). b) Plots showing a positive correlation at the within-individual level (rmcorr) between the discriminability d’ of familiarity-based memory and contrast estimates in the right TPJa [12]. c) Drawing showing deactivation of the VAN/CON/SN that might induce switching from lateral frontoparietal (or central executive) network activity to default-mode network activity.
According to a combination of functional networks and salience processing, we hypothesized that the EI transition, which is generated by the PreBötC, may deactivate the nodes of the right TPJa, right MFG, and dmPFC (i.e., VAN/CON/SN), leading to the opposite direction of salient stimuli that might recruit the default-mode network switched or initiated from the lateral frontoparietal (or central executive) network (Figure 6c). This is a potential reason healthy volunteers could not temporally update or access information that is usually available in their memory, resulting in extended RT and unsuccessful retrieval [12, 13]. This idea may also be relevant to the results that the timing of button-press responses for cognitive processes was tightly distributed ahead of the EI transition [12], suggesting that participants may involuntarily avoid passing through the EI transition during retrieval due to the difficulty in answering correctly. Elucidation of the detailed functional relevance of breathing to large-scale brain network activity with different task demands is a subject for future work.
6. Conclusion
Breathing is a dynamic action, in which cognitive processes can be coordinated with the respiratory cycle and neural network activity and can be constantly updated during online brain states. However, the process is drastically extended when it spans the EI transition, resulting in a failed process. Emerging evidence assumes that the EI transition might reset novelty processing (i.e., encoding and retrieval) and then immediately start consolidation (and reconsolidation). This perspective suggests that the EI transition may play an important role in a “reset and trigger” process whose related neural substrates may be incorporated in the VAN/CON/SN. Thus, stimulation of these functional networks by breathing manipulation could be applied to improve dysfunction, abnormal activity, and/or hyperactivation of the SN in patients with stress responses and neuropsychiatric disorders. These considerations of “brain-breath” interactions would contribute to a better understanding of interoceptive and cognitive mechanisms that underlie brain-body interactions in health conditions and in neuropsychiatric disorders.
Data Availability
Datasets are openly available.
Conflicts of Interest
None.
Funding
This work was supported by grants from the Hyogo Innovative Challenge, Hyogo Medical University (NHN), Academic Research Grants, Hyogo Science and Technology Association (HyogoSTA, NHN), the Cooperative Study Program of National Institute for Physiological Sciences (NHN), the Takeda Science Foundation (NHN), and the Grant-in-Aid for Scientific Research (15K12055) of the Japan Society for the Promotion of Science (YO).
Acknowledgements
We thank Genevieve Yuen (Weill Cornell Medical College, New York, NY, USA), Yasumasa Okada (Murayama Medical Center, Japan), Norihiro Sadato (National Institute for Physiological Sciences, Okazaki, Japan), and Akari Furuse (Hyogo Medical University, Japan) for comments on and discussion of the manuscript.
REFERENCES
[1] Hugo D Critchley, Neil A Harrison
“Visceral influences on brain and behavior.” Neuron, vol. 77, no. 4, pp.
624-638, 2013. View at: Publisher
Site | PubMed
[2] Karl Friston “The free-energy
principle: a rough guide to the brain?” Trends Cogn Sci, vol. 13, no. 7,
pp. 293-301, 2009. View at: Publisher
Site | PubMed
[3] Anil K Seth “Interoceptive inference,
emotion, and the embodied self.” Trends Cogn Sci, vol. 17, no. 11, pp.
565-573, 2013. View at: Publisher
Site | PubMed
[4] Lisa Feldman Barrett, W Kyle Simmons
“Interoceptive predictions in the brain.” Nat Rev Neurosci, vol. 16, no.
7, pp. 419-429, 2015. View at: Publisher
Site | PubMed
[5] B S McEwen “Protective and damaging
effects of stress mediators.” N Eng J Med, vol. 338, no. 3, pp. 171-179,
1998. View at: Publisher
Site | PubMed
[6] Bruce S McEwen, Huda Akil “Revisiting
the Stress Concept: Implications for Affective Disorders.” J Neurosci,
vol. 40, no. 1, pp. 12-21, 2020. View at: Publisher Site | PubMed
[7] O Devinsky, M J Morrell, B A Vogt
“Contributions of anterior cingulate cortex to behaviour.” Brain, vol.
118, no. 1, pp. 279-306, 1995. View at: Publisher Site | PubMed
[8] Ravinder Jerath, Connor Beveridge
“Respiratory Rhythm, Autonomic Modulation, and the Spectrum of Emotions: The
Future of Emotion Recognition and Modulation.” Front Psychol, vol. 11,
pp. 1980, 2020. View at: Publisher
Site | PubMed
[9] Vojislav Maric, Dhakshin Ramanathan,
Jyoti Mishra “Respiratory regulation & interactions with neuro-cognitive
circuitry.” Neurosci Biobehav Rev, vol. 112, pp. 95-106, 2020. View at: Publisher Site | PubMed
[10] Asena Boyadzhieva, Ezgi Kayhan
“Keeping the Breath in Mind: Respiration, Neural Oscillations, and the Free
Energy Principle.” Front Neurosci, vol. 15, pp. 647579, 2021. View at: Publisher Site | PubMed
[11] Tiina Parviainen, Pessi Lyyra, Miriam
S Nokia “Cardiorespiratory rhythms, brain oscillatory activity and cognition:
review of evidence and proposal for significance.” Neurosci Biobehav Rev,
vol. 142, 104908, 2022. View at: Publisher Site | PubMed
[12] Nozomu H Nakamura, Masaki Fukunaga,
Tetsuya Yamamoto, et al. “Respiration-timing-dependent changes in activation of
neural substrates during cognitive processes.” Cereb Cortex Commun, vol.
3, no, 4, pp. tgac038, 2022. View at: Publisher Site | PubMed
[13] Nozomu H Nakamura, Masaki Fukunaga,
Yoshitaka Oku “Respiratory modulation of cognitive performance during the
retrieval process.” PLoS One, vol. 13, no. 9, pp. e0204021, 2018. View
at: Publisher
Site | PubMed
[14] Maurizio Corbetta, Gaurav Patel,
Gordon L Shulman “The Reorienting System of the Human Brain: From Environment
to Theory of Mind.” Neuron, vol. 58, no. 3, pp. 306-324, 2008. View at: Publisher Site | PubMed
[15] Kajsa M Igelström, Michael S A
Graziano “The inferior parietal lobule and temporoparietal junction: A network
perspective.” Neuropsychologia, vol. 105, pp. 70-83, 2017. View at: Publisher
Site | PubMed
[16] Hanaan Bing-Canar, Jacquelyne
Pizzuto, Rebecca J Compton “Mindfulness-of-breathing exercise modulates EEG
alpha activity during cognitive performance.” Psychophysiology, vol. 53,
no. 9, pp. 1366-1376, 2016. View at: Publisher Site | PubMed
[17] Thomas E Dick, Joseph R Mims,
Yee-Hsee Hsieh, et al. “Increased cardio-respiratory coupling evoked by slow
deep breathing can persist in normal humans.” Respir Physiol Neurobiol,
vol. 204, pp. 99-111, 2014. View at: Publisher Site | PubMed
[18] Guy William Fincham, Clara Strauss,
Jesus Montero-Marin, et al. “Effect of breathwork on stress and mental health:
A meta-analysis of randomised-controlled trials.” Sci Rep, vol. 13, no.
1, pp. 432, 2023. View at: Publisher
Site | PubMed
[19] Goldy Yadav, Pratik K Mutha “Deep
Breathing Practice Facilitates Retention of Newly Learned Motor Skills.” Sci
Rep, vol. 6, pp. 37069, 2016. View at: Publisher Site | PubMed
[20] Richard P Brown, Patricia L Gerbarg
“Yoga breathing, meditation, and longevity.” Ann NY Acad Sci, vol. 1172,
pp. 54-62, 2009. View at: Publisher Site | PubMed
[21] Richard P Brown, Patricia L Gerbarg,
Fred Muench “Breathing Practices for Treatment of Psychiatric and
Stress-Related Medical Conditions.” Psychiatr Clin N Am, vol. 36, no. 1,
pp. 121-140, 2013. View at: Publisher
Site | PubMed
[22] Akira Yoshikawa, Yuri Masaoka, Masaki
Yoshida, et al. “Heart Rate and Respiration Affect the Functional Connectivity
of Default Mode Network in Resting-State Functional Magnetic Resonance
Imaging.” Front Neurosci, vol. 14, pp. 631, 2020. View at: Publisher Site | PubMed
[23] Daniel S Kluger, Joachim Gross
“Respiration modulates oscillatory neural network activity at rest.” PLoS
Biol, vol. 19, no. 11, pp. e3001457, 2021. View at: Publisher Site | PubMed
[24] Jose L Herrero, Simon Khuvis, Erin
Yeagle, et al. “Breathing above the brain stem: volitional control and
attentional modulation in humans.” J Neurophysiol, vol. 119, no. 1, pp.
145-159, 2018. View at: Publisher
Site | PubMed
[25] Xingchao Wang, Qiong Wu, Laura Egan,
et al. “Anterior insular cortex plays a critical role in interoceptive
attention.” Elife, vol. 8, pp. e42265, 2019. View at: Publisher Site | PubMed
[26] Wen G Chen, Dana Schloesser, Angela M
Arensdorf, et al. “The Emerging Science of Interoception: Sensing, Integrating,
Interpreting, and Regulating Signals within the Self.” Trends Neurosci,
vol. 44, no. 1, pp. 3-16, 2021. View at: Publisher Site | PubMed
[27] A D Craig “Interoception: The sense
of the physiological condition of the body.” Curr Opin Neurobiol, vol.
13, no. 4, pp. 500-505. View at: Publisher Site | PubMed
[28] Henry C Evrard “The organization of
the primate insular cortex.” Front Neuroanat, vol. 13, pp. 43, 2019.
View at: Publisher
Site | PubMed
[29] “Dyspnea. Mechanisms, assessment, and
management: a consensus statement. American Thoracic Society” Am. J Respir
Crit Care Med, vol. 159, no. 1, pp. 321-340, 1999. View at: Publisher Site | PubMed
[30] Isato Fukushi, Mieczyslaw Pokorski,
Yasumasa Okada “Mechanisms underlying the sensation of dyspnea.” Respir
Investig, vol. 59, no. 1, pp. 66-80, 2021. View at: Publisher Site | PubMed
[31] Mark B Parshall, Richard M
Schwartzstein, Lewis Adams et al. “An official American thoracic society
statement: Update on the mechanisms, assessment, and management of dyspnea.” Am
J Respir Crit Care Med, vol. 185, no. 4, pp. 435-452, 2012. View at: Publisher Site | PubMed
[32] Daniela Schön, Michael Rosenkranz,
Jan Regelsberger, et al. “Reduced perception of dyspnea and pain after right
insular cortex lesions.” Am J Respir Crit Care Med, vol. 178, no. 11,
pp. 1173-1179, 2008. View at: Publisher Site | PubMed
[33] Mahlega S Hassanpour, W Kyle Simmons,
Justin S Feinstein, et al. “The insular cortex dynamically maps changes in
cardiorespiratory interoception.” Neuropsychopharmacology, 43, 426-434,
2018. View at: Publisher
Site | PubMed
[34] Andreas von Leupoldt, Tobias Sommer,
Sarah Kegat, Hans Jörg Baumann, et al., “The unpleasantness of perceived
dyspnea is processed in the anterior insula and amygdala.” Am J Respir Crit
Care Med, vol. 177, no. 9, pp. 1026-1032, 2008. View at: Publisher Site | PubMed
[35] Peter J. Lang, Margaret M. Bradley,
Bruce N. Cuthbert “International Affective Picture System (IAPS): Affective
Ratings of Pictures and Instruction Manual.” Technical Report A-6, University
of Florida, Florida, 2005.
[36] Rie Lambaek Mikkelsen, Thomas
Middelboe, Charlotta Pisinger, et al. “Anxiety and depression in patients with
chronic obstructive pulmonary disease (COPD). A review.” Nord J Psychiatry,
vol. 58, no. 1, pp. 65-70, 2004. View at: Publisher Site | PubMed
[37] Marsus I Pumar, Curt R Gray, James R
Walsh, et al. “Anxiety and depression-Important psychological comorbidities of
COPD.” J Thorac Dis, vol. 6, no. 11, pp. 1615-1631, 2014. View at: Publisher
Site | PubMed
[38] Rafael C Freire, Giampaolo Perna,
Antonio E Nardi “Panic disorder respiratory subtype: Psychopathology,
laboratory challenge tests, and response to treatment.” Harv Rev Psychiatry,
vol. 18, no. 4, pp. 220-229, 2010. View at: Publisher Site | PubMed
[39] Carolin Wolters, Alexander L Gerlach,
Anna Pohl “Interoceptive accuracy and bias in somatic symptom disorder, illness
anxiety disorder, and functional syndromes: A systematic review and
meta-analysis.” PLoS One, vol. 17, no. 8, pp. e0271717, 2022. View at: Publisher Site | PubMed
[40] Naoshige Uchida, Zachary F Mainen
“Speed and accuracy of olfactory discrimination in the rat.” Nat Neurosci,
vol. 6, no. 11, pp. 1224-1229, 2003. View at: Publisher Site | PubMed
[41] Kevin M Cury, Naoshige Uchida “Robust
Odor Coding via Inhalation-Coupled Transient Activity in the Mammalian
Olfactory Bulb.” Neuron, vol. 68, no. 3, pp. 570-585, 2010. View at: Publisher Site | PubMed
[42] Roman Shusterman, Matthew C Smear,
Alexei A Koulakov “Precise olfactory responses tile the sniff cycle.” Nat
Neurosci, 14, 1039-1044, 2011. View at: Publisher Site | PubMed
[43] John C Curtis, David Kleinfeld
“Phase-to-rate transformations encode touch in cortical neurons of a scanning
sensorimotor system.” Nat Neurosci, vol. 12, no. 4, pp. 492-501, 2009.
View at: Publisher Site | PubMed
[44] David Kleinfeld, Martin Deschênes,
Fan Wang, et al. “More than a rhythm of life: Breathing as a binder of
orofacial sensation.” Nat Neurosci, 17, 647-651, 2014. View at: Publisher Site | PubMed
[45] Yuri Masaoka, Nobuyoshi Koiwa, Ikuo
Homma “Inspiratory phase-locked alpha oscillation in human olfaction: Source
generators estimated by a dipole tracing method.” J Physiol, vol. 566,
no. 3, pp. 979-997, 2005. View at: Publisher Site | PubMed
[46] Martin Grund, Esra Al, Marc Pabst, et
al. “Respiration, Heartbeat, and Conscious Tactile Perception.” J Neurosci,
vol. 42, no. 4, pp. 643-656, 2022. View at: Publisher Site | PubMed
[47] Jerry E. Flexman, Robert G. Demaree,
D. Dwayne Simpson “Respiratory phase and visual signal detection.” Percept
Psychophys, vol. 16, pp. 337-339, 1974. View at: Publisher Site
[48] Daniel S Kluger, Elio Balestrieri,
Niko A Busch et al. “Respiration aligns perception with neural excitability.” Elife,
vol. 10, pp. e70907, 2021. View at: Publisher Site | PubMed
[49] Detlef H Heck, Robert Kozma, Leslie M
Kay “The rhythm of memory: how breathing shapes memory function.” J
Neurophysiol, vol. 122, no. 2, pp. 563-571, 2019. View at: Publisher Site | PubMed
[50] Adriano B L Tort, Jurij Brankačk, Andreas Draguhn “Respiration-entrained brain rhythms
are global but often overlooked.” Trends Neurosci, vol. 41, no. 4, pp. 186-197,
2018. View at: Publisher
Site | PubMed
[51] Alfredo Fontanini, PierFranco Spano,
James M Bower “Ketamine-Xylazine-induced slow (1.5 Hz) oscillations in the rat
piriform (olfactory) cortex are functionally correlated with respiration.” J
Neurosci, vol. 23, no. 22, pp. 7993-8001, 2003. View at: Publisher
Site | PubMed
[52] J Ito, S Roy, Y Liu et al. “Whisker
barrel cortex delta oscillations and gamma power in the awake mouse are linked
to respiration.” Nat Commun, vol. 5, pp. 3572, 2014. View at: Publisher Site | PubMed
[53] Vivan Nguyen Chi, Carola Müller,
Thérèse Wolfenstetter “Hippocampal respiration-driven rhythm distinct from
theta oscillations in awake mice.” J Neurosci, vol. 36, no. 1, pp.
162-177, 2016. View at: Publisher
Site | PubMed
[54] Yu Liu, Samuel S McAfee, Detlef H
Heck “Hippocampal sharp-wave ripples in awake mice are entrained by
respiration.” Sci Rep, vol. 7, no. 1, pp. 8950, 2017. View at: Publisher Site | PubMed
[55] André L V Lockmann, Diego A Laplagne,
Richardson N Leão, et al. “A respiration-coupled rhythm in the rat hippocampus
independent of theta and slow oscillations.” J Neurosci, vol. 36, no.
19, pp. 5338-5352, 2016. View at: Publisher Site | PubMed
[56] Yevgenij Yanovsky, Mareva Ciatipis,
Andreas Draguhn, et al. “Slow oscillations in the mouse hippocampus entrained
by nasal respiration.” J Neurosci, vol. 34, no. 17, pp. 5949-5964. View
at: Publisher
Site | PubMed
[57] Jonatan Biskamp, Marlene Bartos,
Jonas-Frederic Sauer “Organization of prefrontal network activity by
respiration-related oscillations.” Sci Rep, vol. 7, pp. 45508, 2017.
View at: Publisher Site | PubMed
[58] Weiwei Zhong, Mareva Ciatipis,
Thérèse Wolfenstetter, et al. “Selective entrainment of gamma subbands by
different slow network oscillations.” Proc Natl Acad Sci U S A, 114,
4519-4524, 2017. View at: Publisher
Site | PubMed
[59] Christina Zelano, Heidi Jiang,
Guangyu Zhou “Nasal respiration entrains human limbic oscillations and
modulates cognitive function.” J Neurosci, vol. 36, no. 49, pp.
12448-12467, 2016. View at: Publisher Site | PubMed
[60] Erik W Schomburg, Antonio
Fernández-Ruiz, Kenji Mizuseki, et al. “Theta Phase Segregation of
Input-Specific Gamma Patterns in Entorhinal-Hippocampal Networks.” Neuron,
84, 470-485, 2014. View at: Publisher Site | PubMed
[61] Makoto Tamura, Timothy J Spellman,
Andrew M Rosen, et al. “Hippocampal-prefrontal theta-gamma coupling during
performance of a spatial working memory task.” Nat Commun, vol. 8, no.
1, pp. 2182, 2017. View at: Publisher
Site | PubMed
[62] Adriano B L Tort, Robert W
Komorowski, Joseph R Manns, et al. “Theta-gamma coupling increases during the
learning of item-context associations.” Proc Natl Acad Sci U S A, vol.
106, no. 49, pp. 20942-20947, 2009. View at: Publisher Site | PubMed
[63] Nikolai Axmacher, Melanie M Henseler,
Ole Jensen, et al. “Cross-frequency coupling supports multi-item working memory
in the human hippocampus.” Proc. Natl. Acad Sci U S A, vol. 107, no. 7,
pp. 3228-3233, 2010. View at: Publisher
Site | PubMed
[64] Uwe Friese, Moritz Köster, Uwe
Hassler, et al. “Successful memory encoding is associated with increased
cross-frequency coupling between frontal theta and posterior gamma oscillations
in human scalp-recorded EEG.” Neuroimage, voll. 66, pp. 642-647, 2013.
View at: Publisher
Site | PubMed
[65] Moritz Köster, Uwe Friese, Benjamin
Schöne, et al. “Theta-gamma coupling during episodic retrieval in the human
EEG.” Brain Res, vol. 1577, pp. 57-68, 2014. View at: Publisher Site | PubMed
[66] R V Baudinette, B J Gannon, W B
Runciman, et al. “Do Cardiorespiratory frequencies show entrainment with
hopping in the tammar wallaby?” J Exp Biol, vol. 129, pp. 251-263, 1987.
View at: Publisher
Site | PubMed
[67] Roger E. Carpenter “Flight physiology
of intermediate-sized fruit bats (Pteropodidae).” J Exp Biol, vol. 120,
pp. 79-103, 1986. View at: Publisher
Site
[68] D M Bramble, F A Jenkins Jr
“Mammalian Locomotor-Respiratory Integration: Implications for Diaphragmatic
and Pulmonary Design.” Science, vol. 262, no. 5131, pp. 235-240, 1993.
View at: Publisher
Site | PubMed
[69] Eric M Lamberg, Jason H Mateika,
Lance Cherry “Internal representations underlying respiration during object
manipulation.” Brain Res, vol. 982, no. 2, pp. 270-279, 2003. View at: Publisher Site | PubMed
[70] J H Mateika, A M Gordon “Adaptive and
dynamic control of respiratory and motor systems during object manipulation.” Brain
Res, vol. 864, no. 2, pp. 327-337, 2000. View at: Publisher Site | PubMed
[71] Hyeong-Dong Park, Coline Barnoud,
Henri Trang, et al. “Breathing is coupled with voluntary action and the
cortical readiness potential.” Nat Commun, vol. 11, pp. 289, 2020. View
at: Publisher
Site | PubMed
[72] Michelle Johannknecht, Christoph
Kayser “The influence of the respiratory cycle on reaction times in
sensory-cognitive paradigms.” Sci Rep, vol. 12, no. 1, pp. 2586, 2022.
View at: Publisher
Site | PubMed
[73] Artin Arshamian, Behzad Iravani,
Asifa Majid “Respiration modulates olfactory memory consolidation in humans. J.
Neurosci, vol. 38, no. 48, pp. 10286-10294, 2018. View at: Publisher Site | PubMed
[74] Tomi Waselius, Jan Wikgren, Markku
Penttonen, et al. “Breathe out and learn: Expiration-contingent stimulus
presentation facilitates associative learning in trace eyeblink conditioning.” Psychophysiology,
vol. 56, no. 9, pp. e13387, 2019. View at: Publisher Site | PubMed
[75] Nicola S Clayton, Timothy J Bussey,
Anthony Dickinson “Can animals recall the past and plan for the future?” Nat
Rev Neurosci, vol. 4, no. 8, pp. 685-691, 2003. View at: Publisher Site | PubMed
[76] H Eichenbaum, A P Yonelinas, C
Ranganath “The medial temporal lobe and recognition memory.” Annu Rev
Neurosci, vol. 30, pp. 123-152, 2007. View at: Publisher
Site | PubMed
[77] Gardiner J. M. “Retrieval: On its
essence and related concepts. In: Science of Memory: Concepts” (Roediger HL,
Dudai Y, Fitzpatrick SM ed), New York: Oxford University Press. pp.221-224,
2007.
[78] Diethelm W Richter, Jeffrey C Smith
“Respiratory rhythm generation in vivo.” Physiology (Bethesda), vol. 29,
no. 1, pp. 58-71, 2014. View at: Publisher Site | PubMed
[79] Jeffrey C Smith, Ana P L Abdala, Anke
Borgmann, et al. “Brainstem respiratory networks: Building blocks and
microcircuits.” Trends Neurosci, vol. 36, no. 3, pp. 152-162, 2013. View
at: Publisher
Site | PubMed
[80] Ofer Perl, Aharon Ravia, Mica
Rubinson, et al. “Human non-olfactory cognition phase-locked with inhalation.” Nat
Hum Behav, vol. 3, no. 5, pp. 501-512, 2019. View at: Publisher Site | PubMed
[81] J C Smith, H H Ellenberger, K
Ballanyi, et al. “Pre-Bötzinger complex: A brainstem region that may generate
respiratory rhythm in mannals.” Science, vol. 254, no. 5032, pp.
726-729, 1991. View at: Publisher
Site | PubMed
[82] Hiroshi Onimaru, Ikuo Homma “A novel
functional neuron group for respiratory rhythm generation in the ventral
medulla.” J Neurosci, 23, 1478-1486, 2003. View at: Publisher
Site | PubMed
[83] Robert T R Huckstepp, Kathryn P
Cardoza, Lauren E Henderson, et al. “Role of parafacial nuclei in control of
breathing in adult rats.” J Neurosci, vol. 35, no. 3, pp. 1052-1067,
2015. View at: Publisher
Site | PubMed
[84] Wiktor A Janczewski, Jack L Feldman
“Distinct rhythm generators for inspiration and expiration in the juvenile
rat.” J Physiol, vol. 570, no. 2, pp. 407-420, 2006. View at: Publisher Site | PubMed
[85] Jack L Feldman, Christopher A Del
Negro, Paul A Gray “Understanding the rhythm of breathing: So near, yet so
far.” Annu Rev Physiol, vol. 75, pp. 423-452, 2013. View at: Publisher
Site | PubMed
[86] Mathias Dutschmann, Thomas E Dick
“Pontine mechanisms of respiratory control.” Compr Physiol, vol. 2, no.
4, pp. 2443-2469, 2012. View at: Publisher
Site | PubMed
[87] Christopher A Del Negro, Gregory D
Funk, Jack L Feldman “Breathing matters.” Nat Rev Neurosci, vol. 19, no.
6, pp. 351-367, 2018. View at: Publisher Site | PubMed
[88] Hidehiko Koizumi, Naohiro Koshiya,
Justine X Chia, et al. “Structural-functional properties of identified
excitatory and inhibitory interneurons within pre-Bötzinger complex respiratory
microcircuits.” J Neurosci, vol. 33, no. 7, pp. 2994-3009, 2013. View
at: Publisher
Site | PubMed
[89] N Koshiya, Y Oku 2, S Yokota, et al.
“Anatomical and functional pathways of rhythmogenic inspiratory premotor
information flow originating in the pre-Bötzinger complex in the rat medulla.” Neuroscience,
vol. 268, pp. 194-211, 2014. View at: Publisher
Site | PubMed
[90] N Koshiya, J C Smith “Neuronal
pacemaker for breathing visualized in vitro.” Nature, vol. 400, no.
6742, 99. 360-363, 1999. View at: Publisher
Site | PubMed
[91] Leszek Kubin, George F Alheid, Edward
J Zuperku, et al. “Central pathways of pulmonary and lower airway vagal
afferents.” J Appl Physiol, vol. 101, no. 2, pp. 618-627, 2006. View at:
Publisher
Site | PubMed
[92] K Ezure, I Tanaka “Distribution and
medullary projection of respiratory neurons in the dorsolateral pons of the
rat.” Neuroscience, vol. 141, no. 2, pp. 1011-1023, 2006. View at: Publisher
Site | PubMed
[93] Tatiana M Anderson, Alfredo J Garcia
3rd, Nathan A Baertsch. “A novel excitatory network for the control of
breathing.” Nature, vol. 536, no. 7614, pp. 76-80, 2016. View at: Publisher Site | PubMed
[94] Jan-Marino Ramirez, Nathan A Baertsch
“The dynamic basis of respiratory rhythm generation: One breath at a time.” Annu
Rev Neurosci, vol. 41, pp. 475-499, 2018. View at: Publisher
Site | PubMed
[95] Rahat Ul Ain Summan Toor, Qi-Jian
Sun, Natasha N Kumar, et al. “Neurons in the intermediate reticular nucleus
coordinate postinspiratory activity, swallowing, and respiratory-sympathetic
coupling in the rat.” J Neurosci, vol. 39, no. 49, pp. 9757-9766, 2019.
View at: Publisher
Site | PubMed
[96] J C Smith, A P L Abdala, H Koizumi,
et al. “Spatial and functional architecture of the mammalian brain stem
respiratory network: A hierarchy of three oscillatory mechanisms.” J
Neurophysiol, vol. 98, no. 6, pp. 3370-3387, 2007. View at: Publisher Site | PubMed
[97] Sarah F Hassan, Jennifer L Cornish, Ann
K Goodchild “Respiratory, metabolic and cardiac functions are altered by
disinhibition of subregions of the medial prefrontal cortex.” J Physiol,
vol. 591, no. 23, pp. 6069-6088, 2013. View at: Publisher Site | PubMed
[98] S C Ramsay, L Adams, K Murphy, et al.
“Regional cerebral blood flow during volitional expiration in man: a comparison
with volitional inspiration.” J Physiol, vol. 461, pp. 85-101, 1993.
View at: Publisher
Site | PubMed
[99] Jonathan D Power, Charles J Lynch,
Marc J Dubin, et al. “Characteristics of respiratory measures in young adults
scanned at rest, including systematic changes and “missed” deep breaths.” Neuroimage,
vol. 204, pp. 116234, 2020. View at: Publisher Site | PubMed
[100] S Ogawa, T M Lee, A R Kay, et al.
“Brain magnetic resonance imaging with contrast dependent on blood
oxygenation.” Proc Natl Acad Sci U S A, vol. 87, no. 24, pp. 9868-9872,
1990. View at: Publisher
Site | PubMed
[101] Seong-Gi Kim, Seiji Ogawa
“Biophysical and physiological origins of blood oxygenation level-dependent
fMRI signals.” J Cereb Blood Flow Metab, vol. 32, no. 7, pp. 1188-1206,
2012. View at: Publisher
Site | PubMed
[102] Kevin Murphy, Rasmus M Birn, Peter A
Bandettini “Resting-state fMRI confounds and cleanup.” Neuroimage, vol.
80, pp. 349-359, 2013. View at: Publisher Site | PubMed
[103] N K Logothetis, J Pauls, M Augath, et
al. “Neurophysiological investigation of the basis of the fMRI signal.” Nature,
vol. 412, no. 6843, pp. 150-157, 2001. View at: Publisher Site | PubMed
[104] Ahalya Viswanathan, Ralph D Freeman
“Neurometabolic coupling in cerebral cortex reflects synaptic more than spiking
activity.” Nat Neurosci, vol. 10, no. 10, pp. 1308-1312, 2007. View at: Publisher Site | PubMed
[105] Costantino Iadecola, Maiken
Nedergaard “Glial regulation of the cerebral microvasculature.” Nat Neurosci,
vol. 10, no. 11, pp. 1369-1376, 2007. View at: Publisher Site | PubMed
[106] Rasmus M Birn, Jason B Diamond,
Monica A Smith, et al. “Separating respiratory-variation-related fluctuations
from neuronal-activity-related fluctuations in fMRI.” Neuroimage, vol.
31, no. 4, pp. 1536-1548, 2006. View at: Publisher Site | PubMed
[107] Rasmus M Birn “The role of
physiological noise in resting-state functional connectivity.” Neuroimage,
vol. 62, no. 2, pp. 864-870, 2012. View at: Publisher Site | PubMed
[108] Willem Huijbers, Cyriel M A Pennartz,
Ewa Beldzik, et al. “Respiration phase-locks to fast stimulus presentations:
Implications for the interpretation of posterior midline “deactivations.”” Hum
Brain Mapp, vol. 35, no. 9, pp. 4932-4943, 2014. View at: Publisher Site | PubMed
[109] Rasmus M Birn, Kevin Murphy, Daniel A
Handwerker, et al. “fMRI in the presence of task-correlated breathing
variations.” Neuroimage, vol. 47, no. 3, pp. 1092-1104, 2009. View at: Publisher Site | PubMed
[110] Rasmus M Birn, Monica A Smith, Tyler
B Jones, et al. “The respiration response function: The temporal dynamics of
fMRI signal fluctuations related to changes in respiration.” Neuroimage,
vol. 40, no. 2, pp. 644-654, 2008. View at: Publisher Site | PubMed
[111] K. J. Friston, P. Jezzard, R. Turner
“Analysis of functional MRI time‐series.” Hum Brain Mapp, vol. 1, no.
2, pp. 153-171, 1994. View at: Publisher
Site
[112] K J Worsley, K J Friston “Analysis of
fMRI time-series revisited--again.” Neuroimage, vol. 2, no. 3, pp.
173-181, 1995. View at: Publisher
Site | PubMed
[113] Ludovica Griffanti, Gwenaëlle Douaud,
Janine Bijsterbosch, et al. “Hand classification of fMRI ICA noise components.”
Neuroimage, vol. 154, pp. 188-205, 2017. View at: Publisher Site | PubMed
[114] Yoshihisa Okamoto; Takafumi Sasaoka;
Norihiro Sadato, et al. “Is Human Brain Activity during Driving Operations
Modulated by the Viscoelastic Characteristics of a Steering Wheel?: An fMRI
Study.” IEEE Access, vol. 8, pp. 215073-215090, 2020. View at: Publisher Site
[115] Ludovica Griffanti, Gholamreza
Salimi-Khorshidi, Christian F Beckmann, et al. “ICA-based artefact removal and
accelerated fMRI acquisition for improved resting state network imaging.” Neuroimage,
vol. 95, pp. 232-247, 2014. View at: Publisher Site | PubMed
[116] Cindy F Yang 1, Jack L Feldman
“Efferent projections of excitatory and inhibitory preBötzinger Complex
neurons.” J Comp Neurol, vol. 526, no. 8, pp. 1389-1402, 2018. View at: Publisher Site | PubMed
[117] Patrick M Fuller, David Sherman,
Nigel P Pedersen, et al. “Reassessment of the structural basis of the ascending
arousal system.” J Comp Neurol, 519, 933-956, 2011. View at: Publisher Site | PubMed
[118] Kevin Yackle, Lindsay A Schwarz,
Kaiwen Kam, et al. “Breathing control center neurons that promote arousal in
mice.” Science, vol. 355, no. 6332, pp. 1411-1415, 2017. View at: Publisher Site | PubMed
[119] Gary Aston-Jones, Jonathan D Cohen
“An integrative theory of locus coeruleus-norepinephrine function: Adaptive
gain and optimal performance.” Annu Rev Neurosci, vol, 28, pp. 403-450,
2005. View at: Publisher
Site | PubMed
[120] Sander Nieuwenhuis, Gary Aston-Jones,
Jonathan D Cohen “Decision making, the P3, and the locus
coeruleus-norepinephrine system.” Psychol Bull, vol. 131, no. 4, pp.
510-532, 2005. View at: Publisher
Site | PubMed
[121] Maurizio Corbetta, Gordon L Shulman
“Control of goal-directed and stimulus-driven attention in the brain.” Nat
Rev Neurosci, vol. 3, no. 3, pp. 201-215, 2002. View at: Publisher Site | PubMed
[122] Gina R Poe, Stephen Foote, Oxana
Eschenko, et al. “Locus coeruleus: a new look at the blue spot.” Nat Rev
Neurosci, vol. 21, no. 11, pp. 644-659, 2020. View at: Publisher Site | PubMed
[123] Susan J Sara, Sebastien Bouret
“Orienting and reorienting: The locus coeruleus mediates cognition through
arousal.” Neuron, vol. 76, no. 1, pp. 130-141, 2012. View at: Publisher Site | PubMed
[124] Elena M Vazey, David E Moorman, Gary
Aston-Jones “Phasic locus coeruleus activity regulates cortical encoding of
salience information.” Proc Natl Acad Sci U S A, vol. 115, no. 40, pp.
E9439-E9448, 2018. View at: Publisher
Site | PubMed
[125] Rogier B Mars, Jérôme Sallet, Urs
Schüffelgen, et al. “Connectivity-based subdivisions of the human right
“temporoparietal junction area”: Evidence for different areas participating in
different cortical networks.” Cereb Cortex, vol. 22, no. 8, pp.
1894-1903, 2012. View at: Publisher
Site | PubMed
[126] Kajsa M Igelström, Taylor W Webb,
Michael S A Graziano “Neural processes in the human temporoparietal cortex
separated by localized independent component analysis.” J Neurosci, vol.
35, no. 25, pp. 9432-9445, 2015. View at: Publisher Site | PubMed
[127] Christopher L Asplund, J Jay Todd,
Andy P Snyder, et al. “A central role for the lateral prefrontal cortex in
goal-directed and stimulus-driven attention.” Nat. Neurosci, vol. 13,
no. 4, pp. 507-512, 2010. View at: Publisher
Site | PubMed
[128] Vinod Menon, Lucina Q Uddin
“Saliency, switching, attention and control: a network model of insula
function.” Brain Struct Funct, vol. 214, no. 5-6, pp. 655-667, 2010.
View at: Publisher
Site | PubMed
[129] B T Thomas Yeo, Fenna M Krienen,
Jorge Sepulcre, “The organization of the human cerebral cortex estimated by
intrinsic functional connectivity.” J Neurophysiol, vol. 106, no. 3, pp.
1125-1165, 2011. View at: Publisher
Site | PubMed
[130] Martijn P van den Heuvel, Olaf Sporns
“An anatomical substrate for integration among functional networks in human
cortex.” J Neurosci, vol. 33, no. 36, pp. 14489-14500, 2013. View at: Publisher Site | PubMed
[131] Lucina Q Uddin, B T Thomas Yeo, R
Nathan Spreng “Towards a universal taxonomy of macro-scale functional human
brain networks.” Brain Topogr, vol. 32, no. 6, pp. 926-942, 2019. View at:
Publisher Site | PubMed
[132] Rogier B Mars, Stefan Debener, Thomas
E Gladwin, et al. “Trial-by-trial fluctuations in the event-related
electroencephalogram reflect dynamic changes in the degree of surprise.” J
Neurosci, vol. 28, no. 47, pp. 12539-12545, 2008. View at: Publisher Site | PubMed
[133] Joy J Geng, Simone Vossel
“Re-evaluating the role of TPJ in attentional control: Contextual updating?” Neurosci
Biobehav Rev, vol. 37, no, 10, pp. 2608-2620, 2013. View at: Publisher Site | PubMed
[134] Sander Nieuwenhuis, Eco J De Geus,
Gary Aston-Jones “The anatomical and functional relationship between the P3 and
autonomic components of the orienting response.” Psychophysiology, vol.
48, no. 2, pp. 162-175, 2011. View at: Publisher Site | PubMed
[135] Bryan A Strange, Raymond J Dolan
“Beta-adrenergic modulation of oddball responses in humans.” Behav Brain
Funct, vol. 3, pp. 29, 2007. View at: Publisher Site | PubMed
[136] Thomas Liebe, Jörn Kaufmann, Meng Li,
et al. “In vivo anatomical mapping of human locus coeruleus functional
connectivity at 3 T MRI.” Hum Brain Mapp, vol. 41, no. 8, pp. 2136-2151,
2020. View at: Publisher
Site | PubMed
[137] Thomas Liebe, Milos Dordevic, Jörn
Kaufmann, et al. “Investigation of the functional pathogenesis of mild
cognitive impairment by localisation-based locus coeruleus resting-state fMRI.”
Hum Brain Mapp, vol. 43, no. 18, pp. 5630-5642, 2022. View at: Publisher Site | PubMed
[138] Laurel S Morris, Jordan G McCall 2,
Dennis S Charney, et al. “The role of the locus coeruleus in the generation of
pathological anxiety.” Brain Neurosci Adv, vol. 4, pp. 1-18, 2020. View
at: Publisher Site | PubMed
[139] Christoph Naegeli, Thomas Zeffiro,
Marco Piccirelli, et al. “Locus Coeruleus Activity Mediates Hyperresponsiveness
in Posttraumatic Stress Disorder.” Biol Psychiatry, vol. 83, no. 3, pp.
254-262, 2018. View at: Publisher
Site | PubMed
[140] Vinod Menon “Large-scale brain
networks and psychopathology: A unifying triple network model.” Trends Cogn
Sci, vol. 15, no. 10, pp. 483-506, 2011. View at: Publisher Site | PubMed
[141] William W Seeley 1, Vinod Menon, Alan
F Schatzberg,et al. “Dissociable intrinsic connectivity networks for salience
processing and executive control.” J Neurosci, vol. 27, no. 9, pp.
2349-2356, 2007. View at: Publisher Site | PubMed
[142] Genevieve S Yuen, Faith M
Gunning-Dixon, Matthew J Hoptman, et al. “The salience network in the apathy of
late-life depression.” Int J Geriatr Psychiatry, vol. 29, no. 11, pp.
1116-1124, 2014. View at: Publisher
Site | PubMed
[143] Madeleine Goodkind 1, Simon B
Eickhoff 2, Desmond J Oathes, et al. “Identification of a common
neurobiological substrate for mental Illness.” JAMA Psychiatry, vol. 72,
no. 4, pp. 305-315, 2015. View at: Publisher Site | PubMed
[144] Sarah K Peters, Katharine Dunlop 1,
Jonathan Downar “Cortico-striatal-thalamic loop circuits of the salience
network: A central pathway in psychiatric disease and treatment.” Front Syst
Neurosci, vol. 10, pp. 104, 2016. View at: Publisher
Site | PubMed
[145] Janis Brakowski, Simona Spinelli,
Nadja Dörig, et al. “Resting state brain network function in major depression -
Depression symptomatology, antidepressant treatment effects, future research.” J
Psychiatr Res, View at: Publisher Site | PubMed
[146] Andrei Manoliu, Valentin Riedl,
Andriy Zherdin, et al. “Aberrant dependence of default mode/central executive
network interactions on anterior insular salience network activity in
schizophrenia.” Schizophr Bull, vol. 40, no. 2, pp. 428-437, 2014. View
at: Publisher Site | PubMed
[147] Tereza Nekovarova 1, Iveta Fajnerova
2, Jiri Horacek, et al. “Bridging disparate symptoms of schizophrenia: A triple
network dysfunction theory.” Front Behav Neurosci, vol. 8, pp. 171,
2014. View at: Publisher
Site | PubMed
[148] Daniel C M O'Doherty, Kate M Chitty,
Sonia Saddiqui, et al. “A systematic review and meta-analysis of magnetic
resonance imaging measurement of structural volumes in posttraumatic stress
disorder.” Psychiatry Res, vol. 232, no. 1, pp. 1-33, 2015. View at: Publisher
Site | PubMed
[149] Teddy J Akiki, Christopher L Averill,
Chadi G Abdallah “A Network-Based Neurobiological Model of PTSD: Evidence From
Structural and Functional Neuroimaging Studies.” Curr. Psychiatry Rep,
vol. 19, no. 11, pp. 81, 2017. View at: Publisher Site | PubMed
[150] David J Sharp, Gregory Scott, Robert Leech “Network dysfunction after traumatic brain injury.” Nat Rev Neurol, vol. 10, no. 3, pp. 156-166, 2014. View at: Publisher Site | PubMed
[151] Randall S Scheibel “Functional magnetic resonance imaging of cognitive control following traumatic brain injury.” Front Neurol, vol 8, pp. 352, 2017. View at: Publisher Site | PubMed